Система эпхх: СИСТЕМА УПРАВЛЕНИЯ ЭКОНОМАЙЗЕРОМ ПРИНУДИТЕЛЬНОГО ХОЛОСТОГО ХОДА (ЭПХХ). ОСОБЕННОСТИ КОНСТРУКЦИИ
Система ЭПХХ, схема подключения эпхх, необходимость система ЭПХХ
просмотров 8 614 Google+Система ЭПХХ предназначена для прекращения подачи топлива при следовании автомобиля в режиме принудительного холостого хода. Этот режим характеризуется частотой вращения коленвала двигателя, который превышает обороты холостого хода при закрытой дроссельной заслонке. Такой режим применяется в городском цикле и следовании под гору в режиме торможения двигателем.
На инжекторных двигателях прекращение подачи топлива осуществляет электронная система управления двигателем, а в карбюраторных двигателях блок управления ЭПХХ.
Из чего состоит система ЭПХХ.
В состав система ЭПХХ входит блок управления, электромагнитный клапан или электромагнитный пневмоклапан, датчик положения дроссельной заслонки. В качестве датчика числа оборотов часто используется прерыватель трамблёра.
Датчиком положения дроссельной заслонки может быть микропереключатель, контакты которого размыкаются при закрытой дроссельной заслонке либо датчик-винт на конце, которого крепится провод соединяющий вывод блока управления с массой при открытой дроссельной заслонке.
Датчиком оборотов в карбюраторном двигателе выступает прерыватель трамблёра.
Прекращение подачи топлива осуществляется электромагнитным клапаном или электропневматическим клапаном, в зависимости от конструкции карбюратора. Электромагнитный клапан устанавливается на карбюраторе и перекрывает, при отсутствии питания на нём, своим сердечником канал холостого хода. Электропневматический клапан устанавливается на корпусе автомобиля в разрыве трубки соединяющей впускной коллектор с модулем экономайзера, при включении отключает экономайзер от коллектора и соединяет его с атмосферой.В результате этого клапан экономайзера прекращает подачу топлива.
Принцип работы система ЭПХХ.
При частоте вращения коленвала более 1100 об/мин. и закрытой дроссельной заслонке, блок управления отключает питание с клапана, что прекращает подачу топлива, что позволяет экономить топливо на 2-3% и снизить токсичность выхлопных газов на 15-30%. Кроме этого система ЭПХХ предотвращает детонацию двигателя при отключении двигателя, то есть детонационное сгорание топлива при выключенном зажигании.
Необходимость системы ЭПХХ.
Езда автомобиля с применением принудительного холостого хода происходит крайне редко, в основном в гористой местности. По этому даже обещанные 2-3% практически не достижимая цель. Но предотвращение детонации двигателя это очень часто необходимо. Но для реализации предотвращения детонации при отключении зажигания не обязательно подключения всей схемы. Для этого достаточно просто подать питание на клапан при включении зажигания и снятие его при выключении.
admin 18/06/2011 «Если Вы заметили ошибку в тексте, пожалуйста выделите это место мышкой и нажмите CTRL+ENTER» «Если статья была Вам полезна, поделитесь ссылкой на неё в соцсетях»Экономайзер принудительного холостого хода (ЭПХХ) карбюратора Солекс 2108, 21081, 21083

ЭПХХ является одной из систем карбюратора 2108, 21081, 21083 Солекс. Она отвечает за обеспечение работы двигателя автомобиля на принудительном холостом ходу.
Назначение системы ЭПХХ Солекс
Система экономайзера принудительного холостого хода карбюратора Солекс 2108, 21081, 21083 предназначена для отключения подачи топлива через систему холостого хода на режиме торможения двигателем и после выключения зажигания. Она обеспечивает сокращение расхода топлива до 0,5 литра на 100 км, предотвращает возникновение дизелинга (калильного зажигания), сокращает выброс в атмосферу токсичных соединений образующихся при ухудшении смесеобразования на режиме ПХХ.
Устройство ЭПХХ Солекс
Электромагнитный клапан карбюратора – исполнительное устройство системы, перекрывающее своей иглой, по команде блока управления, отверстие в топливном жиклере системы ХХ, тем самым прекращая через нее подачу топлива. Установлен в карбюраторе.
Электромагнитный клапан карбюратора Солекс 2108, 21081Блок управления – электронный блок управления является управляющим элементом ЭПХХ. Он считывает частоту импульсов в системе зажигания (через вывод «К» катушки зажигания) и по ним определяет частоту вращения коленчатого вала двигателя. Помимо этого блок управления принимает сигнал от контакта винта «количества» топливной смеси о закрытии, либо открытии дроссельной заслонки первой камеры. При определенной частоте вращения к/вала и сигнале об открытой дроссельной заслонки он отключает подачу напряжения на электромагнитный клапан, а тот перекрывает подачу топлива через СХХ. Блок установлен на щитке моторного отсека рядом с коммутатором системы зажигания.
Он считывает частоту импульсов в системе зажигания (через вывод «К» катушки зажигания) и по ним определяет частоту вращения коленчатого вала двигателя. Помимо этого блок управления принимает сигнал от контакта винта «количества» топливной смеси о закрытии, либо открытии дроссельной заслонки первой камеры. При определенной частоте вращения к/вала и сигнале об открытой дроссельной заслонки он отключает подачу напряжения на электромагнитный клапан, а тот перекрывает подачу топлива через СХХ. Блок установлен на щитке моторного отсека рядом с коммутатором системы зажигания.
Контакт винта «количества» топливной смеси (датчик-винт) – наконечник винта «количества» топливной смеси с присоединенным к нему проводом. При отпущенной педали «газа» и закрытых дроссельных заслонках контакт касается ребра рычага на оси дроссельной заслонки (замкнут на «массу»), на блок управления идет сигнал о том, что дроссельная заслонка закрыта.
После нажатия на педаль «газа» контакт наконечника винта «количества» и рычага дроссельной заслонки размыкается (не замкнут на «массу»), на блок управления идет сигнал, что дроссельная заслонка открыта. Установлен на карбюраторе.
Видимые элементы системы ЭПХХ карбюратора Солекс в подкапотном пространстве автомобиля ВАЗ 21083Принцип действия системы экономайзера принудительного холостого хода карбюратора Солекс
Движение автомобиля по инерции с включенной передачей и отпущенной педалью «газа» (торможение двигателем) называется принудительным холостым ходом (ПХХ). На этом режиме резко ухудшаются условия сгорания топливной смеси в цилиндрах двигателя, увеличивается выброс СО и СН, возрастает расход топлива. ЭПХХ отключает подачу топлива на режиме ПХХ в цилиндры двигателя, тем самым оптимизируя состав топливной смеси и экономя топливо.
Блок управления в этой ситуации принимает сигналы с замкнувшегося на «массу» контакта на винте «количества» о том, что дроссельная заслонка закрыта и с катушки о частоте вращения коленчатого вала на данный момент. Если частота вращения выше 2100 оборотов в минуту он прекращает подачу напряжения на вывод электромагнитного клапана карбюратора и тот перекроет отверстие топливного жиклера СХХ. Подача топлива через систему холостого хода прекратится. Как только частота вращения коленчатого вала снизится до 1900 об/мин, блок управления возобновит подачу напряжения на электромагнитный клапан. Его игла втянется и откроет отверстие в топливном жиклере СХХ. Система холостого хода заработает.
То есть для принудительного отключения подачи топлива через систему холостого хода электронному блоку управления необходимо два условия – сигнал о закрытой дроссельной заслонке и сигнал о величине оборотов двигателя.
Неисправности в работе двигателя связанные с ЭПХХ Солекс
— Двигатель глохнет в движении автомобиля при сбросе «газа»
Аналогичная неисправность может возникнуть и при засорении системы холостого хода карбюратора. Это нужно учитывать при диагностике неисправностей ЭПХХ.
Это нужно учитывать при диагностике неисправностей ЭПХХ.
— Дизелинг (калильное зажигание)
Двигатель работает некоторое время после выключения зажигания.
Примечания и дополнения
Принудительный холостой ход (ПХХ) – один из режимов работы двигателя. Выполняется при движении автомобиля с включенной передачей и полностью отпущенной педалью «газа». Например, при торможении двигателем или движении на спуске. В этом случае, дроссельные заслонки обеих камер карбюратора полностью закрыты, обороты коленчатого вала двигателя выше 1900 об/мин. Ниже 1900 об/мин вступает в работу система холостого хода.
Дизелинг (калильное зажигание) – непродолжительная работа двигателя после выключения зажигания. Топливо, попадающее в цилиндры двигателя под действием разрежения из карбюратора, воспламеняется от нагретой свечи, происходят вспышки в камерах сгорания, перемещающие поршни. Возможен при неисправной системе ЭПХХ и применении «горячих» свечей (свечей с температурными характеристиками не соответствующими данному типу двигателей).
Еще статьи по карбюраторам Солекс
— Проверка и ремонт системы ЭПХХ карбюратора 2108, 21081, 21083 Солекс
— Неустойчивый холостой ход двигателя с карбюратором Солекс
— Прочистка системы холостого хода карбюратора Солекс
— «Троит» двигатель, причины
— Обороты холостого хода карбюраторного двигателя не поддаются регулировке
Система экономайзера принудительного холостого хода (ЭПХХ)
Система ЭПХХ отключает подачу топлива во впускной трубопровод в режиме торможения двигателем (так называемого принудительного холостого хода), тем самым экономится топливо и уменьшается выброс токсичных веществ в атмосферу.
Система состоит из элктронного блока управления, микропереключателя, клапана экономайзера принудительного холостого хода и электромагнитного клапана (см. схему карбюратора).
Электронный блок управления ЭПХХ (1412.3733 или 25.3761-01) при увеличении оборотов двигателя свыше 1590–1700 мин -1 , отключает питание обмотки электромагнитного клапана. При этом пространство над диафрагмой экономайзера через клапан, соединяется с атмосферой, разряжение падает и клапан экономайзера, под действием пружины перекрывает канал холостого хода. Когда обороты двигателя падают примерно до 1300 мин -1 , блок управления вновь подает напряжение на электромагнитный клапан, он закрывается и в полости над диафрагмой вновь создается разряжение. Клапан экономайзера открывается, обеспечивая доступ топлива в канал холостого хода. Электромагнитный клапан обесточивается также, если выключить зажигание, чем исключается возможность возникновения работы двигателя с самовоспламенением.
При этом пространство над диафрагмой экономайзера через клапан, соединяется с атмосферой, разряжение падает и клапан экономайзера, под действием пружины перекрывает канал холостого хода. Когда обороты двигателя падают примерно до 1300 мин -1 , блок управления вновь подает напряжение на электромагнитный клапан, он закрывается и в полости над диафрагмой вновь создается разряжение. Клапан экономайзера открывается, обеспечивая доступ топлива в канал холостого хода. Электромагнитный клапан обесточивается также, если выключить зажигание, чем исключается возможность возникновения работы двигателя с самовоспламенением.На карбюраторе установлен микровыключатель, который при открытии дроссельной заслонки подает напряжение на электромагнитный клапан напрямую, минуя блок управления.
Снятие и проверка электромагнитного клапан
ПОРЯДОК ВЫПОЛНЕНИЯ
Ослабляем хомуты и снимаем шланги с патрубков клапана, пометив их положение.
При выключенном зажигании отсоединяем провода.
Ключом “на 10” отворачиваем гайку крепления клапана и минусового провода.
Для проверки клапана подуйте в боковую трубку клапана – воздух должен выходить через воздушный фильтр, а при подаче напряжения на контакты – через вторую трубку.
Неисправный клапан заменяем. Устанавливая клапан убедитесь в правильности подсоединения шлангов: боковая трубка соединяется с экономайзером, а центральная – с задроссельным пространством карбюратора.
Проверка и замена блока управления системой ЭПХХ
ПОРЯДОК ВЫПОЛНЕНИЯ
Сдвинув с разъемов электромагнитного клапана изоляцию, подсоединяем вольтметр.
Отсоединяем провода от микропереключателя карбюратора (см. Снятие карбюратора).
Запускаем двигатель. Вольтметр должен показать наличие напряжения. Увеличиваем частоту вращения коленчатого вала двигателя. По достижении 1700 мин -1 блок управления должен отключить электропитание клапана, а после падения частоты вращения до 1300 мин -1 – вновь включить.
Неисправный блок заменяем.
Крестообразной отверткой отворачиваем три самореза и снимаем правую накладку под панелью приборов.
При выключенном зажигании отсоединяем колодку от разъема блока управления.
Отворачиваем винт крепления блока.
Новый блок устанавливаем в обратной последовательности.
Регулировка микропереключателя системы ЭПХХ
ПОРЯДОК ВЫПОЛНЕНИЯ
Отсоединяем провода от микровыключателя (см. Снятие карбюратора).
Подсоединяем к его штекерам омметр.
При отпущенной педали подачи топлива контакты микровыключателя должны быть разомкнуты, при малейшем нажатии на педаль — замкнуты.
Для регулировки момента включения, отверткой ослабляем два винта и, перемещая микропереключатель вдоль овальных отверстий корпуса, добиваемся его оптимального положения.
Эпхх что это такое
Разберем устройство и принцип действия экономайзера принудительного холостого хода (ЭПХХ) карбюраторов семейства Солекс устанавливаемых на двигателя автомобилей ВАЗ 2108, 2109, 21099 и их модификации. ЭПХХ является одной из систем карбюратора 2108, 21081, 21083 Солекс. Она отвечает за обеспечение работы двигателя автомобиля на принудительном холостом ходу.
ЭПХХ является одной из систем карбюратора 2108, 21081, 21083 Солекс. Она отвечает за обеспечение работы двигателя автомобиля на принудительном холостом ходу.
Система экономайзера принудительного холостого хода карбюратора Солекс 2108, 21081, 21083 предназначена для отключения подачи топлива через систему холостого хода на режиме торможения двигателем и после выключения зажигания. Она обеспечивает сокращение расхода топлива до 0,5 литра на 100 км, предотвращает возникновение дизелинга (калильного зажигания), сокращает выброс в атмосферу токсичных соединений образующихся при ухудшении смесеобразования на режиме ПХХ.
Устройство ЭПХХ Солекс
Электромагнитный клапан карбюратора – исполнительное устройство системы, прекрывающее своей иглой, по команде блока управления, отверстие в топливном жиклере системы ХХ, тем самым прекращая через нее подачу топлива. Установлен в карбюраторе.
Блок управления – электронный блок управления является управляющим элементом ЭПХХ.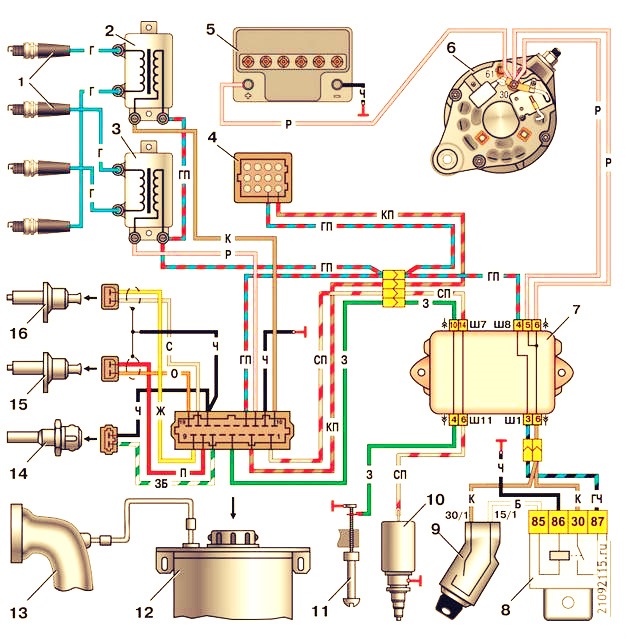 Он считывает частоту импульсов в системе зажигания (через вывод «К» катушки зажигания) и по ним определяет частоту вращения коленчатого вала двигателя. Помимо этого блок управления принимает сигнал от контакта винта «количества» топливной смеси о закрытии, либо открытии дроссельной заслонки первой камеры. При определенной частоте вращения к/вала и сигнале об открытой дроссельной заслонки он отключает подачу напряжения на электромагнитный клапан, а тот перекрывает подачу топлива через СХХ. Блок установлен на щитке моторного отсека рядом с коммутатором системы зажигания.
Он считывает частоту импульсов в системе зажигания (через вывод «К» катушки зажигания) и по ним определяет частоту вращения коленчатого вала двигателя. Помимо этого блок управления принимает сигнал от контакта винта «количества» топливной смеси о закрытии, либо открытии дроссельной заслонки первой камеры. При определенной частоте вращения к/вала и сигнале об открытой дроссельной заслонки он отключает подачу напряжения на электромагнитный клапан, а тот перекрывает подачу топлива через СХХ. Блок установлен на щитке моторного отсека рядом с коммутатором системы зажигания.
Контакт винта «количества» топливной смеси (датчик-винт) – наконечник винта «количества» топливной смеси с присоединенным к нему проводом. При отпущенной педали «газа» и закрытых дроссельных заслонках контакт касается ребра рычага на оси дроссельной заслонки (замкнут на «массу»), на блок управления идет сигнал о том, что дроссельная заслонка закрыта.
После нажатия на педаль «газа» контакт наконечника винта «количества» и рычага дроссельной заслонки размыкается (не замкнут на «массу»), на блок управления идет сигнал, что дроссельная заслонка открыта. Установлен на карбюраторе.
Установлен на карбюраторе.
видимые элементы системы ЭПХХ карбюратора Солекс в подкапотном пространстве автомобиля ВАЗ 21083
Принцип действия системы экономайзера принудительного холостого хода карбюратора Солекс
Движение автомобиля по инерции с включенной передачей и отпущенной педалью «газа» (торможение двигателем) называется принудительным холостым ходом (ПХХ). На этом режиме резко ухудшаются условия сгорания топливной смеси в цилиндрах двигателя, увеличивается выброс СО и СН, возрастает расход топлива. ЭПХХ отключает подачу топлива на режиме ПХХ в цилиндры двигателя, тем самым оптимизируя состав топливной смеси и экономя топливо.
Блок управления в этой ситуации принимает сигналы с замкнувшегося на «массу» контакта на винте «количества» о том, что дроссельная заслонка закрыта и с катушки о частоте вращения коленчатого вала на данный момент. Если частота вращения выше 2100 оборотов в минуту он прекращает подачу напряжения на вывод электромагнитного клапана карбюратора и тот перекроет отверстие топливного жиклера СХХ. Подача топлива через систему холостого хода прекратится. Как только частота вращения коленчатого вала снизится до 1900 об/мин, блок управления возобновит подачу напряжения на электромагнитный клапан. Его игла втянется и откроет отверстие в топливном жиклере СХХ. Система холостого хода заработает.
Подача топлива через систему холостого хода прекратится. Как только частота вращения коленчатого вала снизится до 1900 об/мин, блок управления возобновит подачу напряжения на электромагнитный клапан. Его игла втянется и откроет отверстие в топливном жиклере СХХ. Система холостого хода заработает.
То есть для принудительного отключения подачи топлива через систему холостого хода электронному блоку управления необходимо два условия – сигнал о закрытой дроссельной заслонке и сигнал о величине оборотов двигателя.
Неисправности в работе двигателя связанные с ЭПХХ Солекс
— Двигатель глохнет в движении автомобиля при сбросе «газа»
Аналогичная неисправность может возникнуть и при засорении системы холостого хода карбюратора. Это нужно учитывать при диагностике неисправностей ЭПХХ.
— Дизелинг (калильное зажигание)
Двигатель работает некоторое время после выключения зажигания.
Примечания и дополнения
Принудительный холостой ход (ПХХ) – один из режимов работы двигателя. Выполняется при движении автомобиля с включенной передачей и полностью отпущенной педалью «газа». Например, при торможении двигателем или движении на спуске. В этом случае, дроссельные заслонки обеих камер карбюратора полностью закрыты, обороты коленчатого вала двигателя выше 1900 об/мин. Ниже 1900 об/мин вступает в работу система холостого хода.
Выполняется при движении автомобиля с включенной передачей и полностью отпущенной педалью «газа». Например, при торможении двигателем или движении на спуске. В этом случае, дроссельные заслонки обеих камер карбюратора полностью закрыты, обороты коленчатого вала двигателя выше 1900 об/мин. Ниже 1900 об/мин вступает в работу система холостого хода.
Дизелинг (калильное зажигание) – непродолжительная работа двигателя после выключения зажигания. Топливо, попадающее в цилиндры двигателя под действием разрежения из карбюратора, воспламеняется от нагретой свечи, происходят вспышки в камерах сгорания, перемещающие поршни. Возможен при неисправной системе ЭПХХ и применении «горячих» свечей (свечей с температурными характеристиками не соответствующими данному типу двигателей).
Еще статьи по карбюраторам Солекс
Экономайзер принудительного холостого хода или ЭПХХ позволяет в значительной мере уменьшить выброс токсических веществ в атмосферу. Также он понижает потребление топлива.
Что собой представляет экономайзер
Устройство и схема подключения экономайзера
Устройство ЭПХХ не представляет особой сложности, несмотря на это эффективность системы не поддаётся сомнениям. Стандартная конструкция состоит из таких элементов, как:
- катушка зажигания,
- изолированный наконечник,
- винт,
- электромагнитный клапан,
- блок управления ЭПХХ.
Каждая из этих деталей взаимодействует друг с другом. Результатом подобного процесса является повышенная производительность мотора и существенное увеличение экономии топлива. Но для того чтобы достигнуть такого результата всё должно быть правильно подключено. О том, как это сделать вы можете узнать из схемы подключения ЭПХХ ниже.
Принцип работы
Есть такое понятие, как торможение двигателем. Проще говоря, это ситуация, когда автомобиль продолжает своё движение по инерции. При этом передача всё ещё включена, а педаль, отвечающая за карбюратор отпущена. Подобное состояние также называется принудительным холостым ходом. Отсюда, собственно, и аббревиатура.
Подобное состояние также называется принудительным холостым ходом. Отсюда, собственно, и аббревиатура.
При этом внутри двигателя происходят очень интересные и важные процессы. Естественно, топливная смесь в цилиндрах продолжает воспламеняться. Но при этом эффективность работы системы падает в несколько раз. Как результат отработанные газы имеют повышенное содержание оксида углерода и углеводородов.
Естественно, автомобильные инженеры не могли просто так оставить подобный дефект. Результатом долгих исследований и экспериментов стало изобретение системы ЭПХХ. Она позволяет отключить подачу топлива при работе в режиме холостого хода, тем самым решив ряд описанных выше проблем.
Отключение подачи топлива стало возможным благодаря электромагнитному клапану, который монтируется в крышке карбюратора. В данной конструкции за подачу тока отвечает блок управления. Он вместе с клапаном создаёт одну электрическую цепь, в которую также входит:
- источник питания;
- датчик, фиксирующий положение дроссельной заслонки;
- катушка зажигания,
- масса.


Информация передаётся посредством электрического импульса, который идёт от катушки зажигания. Обычно он содержит данные о частоте вращения. О том, что карбюратор перешёл в режим холостого хода сигнализирует датчик. Это третий контакт, который подключается к одному из винтов. Замыкание делается на массу.
Система ЭПХХ работает таким образом, что на холостом ходу обмотка пятого электромагнитного клапана обесточивается. Результатом этого действия является прекращение подачи топлива.
Для того чтобы подача топлива возобновилась система ЭПХХ при помощи второго блока должна зарегистрировать два изменения:
- Частота вращения коленвала должна превысить отметку в 2000 оборотов за минуту.
- Дроссельная заслонка должна находиться в закрытом положении.
Только тогда, когда эти два условия будут выполнены, система ЭПХХ сможет возобновить подачу топлива. Но не всё так просто. Если с пониманием внутренних процессов никаких сложностей возникнуть не должно, то возникает другой закономерный вопрос, а что для этого нужно сделать водителю?
В действительности всё довольно просто. Чтобы система ЭПХХ возобновила подачу топлива водителю нужно совершить некоторые действия. Вначале необходимо уменьшить скорость движения . При этом нельзя нажимать на педаль, которая контролирует положение дроссельной заслонки.
Чтобы система ЭПХХ возобновила подачу топлива водителю нужно совершить некоторые действия. Вначале необходимо уменьшить скорость движения . При этом нельзя нажимать на педаль, которая контролирует положение дроссельной заслонки.
Дальше вы можете как отжать сцепление, так и перейти на нейтраль. После этих действий режим прекращения подачи топлива будет отключен системой ЭПХХ. Это произойдет из-за нормализации частоты вращений.
Есть ещё один способ дезактивировать систему ЭПХХ. Для этого вам также нужно вдавить в пол педаль, отвечающую за дроссельную заслонку. Но частота вращения должна быть высокой. Чтобы этого добиться нужно продолжать движение.
Отдельно нужно упомянуть особенности работы электромагнитного клапана. Он обесточивается, когда включается зажигание. Подобная предосторожность позволяет исключить то, что двигатель начнёт свою работу с воспламенения.
Неисправности и диагностика ЭПХХ
Система ЭПХХ не отличается особенной сложностью. Именно этот факт выступает гарантией долгой работы. Но даже эта деталь может выйти из строя при больших нагрузках и длительной эксплуатации автомобиля.
Но даже эта деталь может выйти из строя при больших нагрузках и длительной эксплуатации автомобиля.
Обычно при выходе из строя системы — двигатель не запускается при отпущенной педали. Он просто глохнет. Начать диагностику нужно с проверки шланга, который соединяет пневматический электрический клапан и клапан ЭПХХ.
Также нужно при диагностике системы ЭПХХ с большим вниманием отнестись к электрическим контактам. Вы должны проверить надёжность соединений. Довольно часто из строя выходит пневматический электрический клапан. Поэтому очень важно осмотреть и его. Следующими на очереди идут ЭБУ и микропереключатель. Проверку можно проводить только при включённом зажигании и неработающем моторе!
Признаком работоспособности пневматического электроклапана будет характерный щелчок, раздающийся при отсоединении и подключении кабеля. Если же этого нет, то дальнейшую проверку нужно осуществлять при помощи контрольной лампы. Это поможет определить есть ли подача тока. При его отсутствии дальше проверяется ЭБУ и микропереключатель.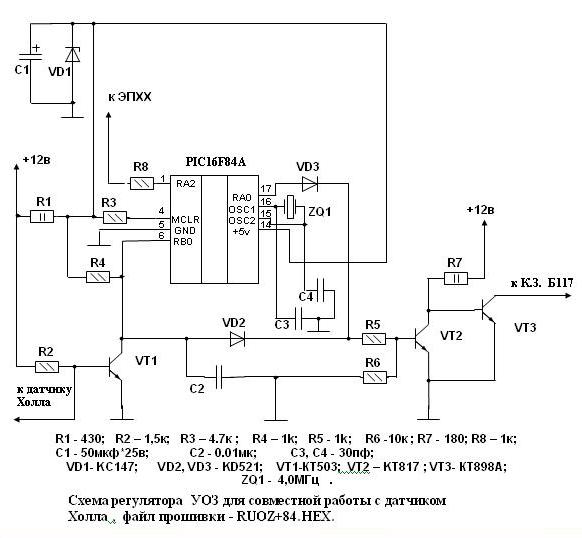
Итоги
ЭПХХ позволяет добиться значительной экономии топлива. Это крайне выгодное конструктивное решение, позволяющее при минимальных затратах повысить производительность мотора. Отдельным бонусом идёт снижение токсичности отработанных газов.
Для управления клапаном ЭПХХ в карбюраторных двигателях автомобилей ВАЗ 2108 — 2110 используется блок управления ЭПХХ 50.3761. В качестве датчика положения дроссельной заслонки используется датчик-винт, представляющий из себя пластмассовый винт с металлическим наконечником, вкручивающийся в кронштейн, закреплённый на карбюраторе.
При открытии дроссельной заслонке наконечник винта, с прикреплённым к нему проводом, не упирается в рычаг дроссельной заслони. Это приводит к разрыву цепи вывода 5 блока управления с массой. При этом закрывается транзистор VT7, а транзистор VT5 открывается, открывая в свою очередь транзистор VT8. Транзистор VT8 подаёт питание на электромагнитный клапан независимо от числа оборотов коленвала.
На вывод 3 блока управления подходит провод, соединяющий его с выводом первичной катушки зажигания, передающий импульсы, которые поступают на вывод 4 микросхемы А1. На выводе 3 микросхемы формируются импульсы постоянной длительности, повторение которых соответствует импульсам от трамблёра. Транзисторы VT1 и VT2 разряжают времязадающий конденсатор С1. Если частота вращения коленвала меньше 1100 об./мин., то напряжение на конденсаторе не поднимается, при повышении числа оборотов напряжение возрастает и когда оно превысит 8 В, происходит открытие транзисторов VT3 и VT4 которые через микросхему А2 открывают транзистор VT6 и соответственно VT8.
Блока управления ЭПХХ 25.3761Схема блока управления ЭПХХ 25.3761 отличается в основном только работой при оборотах коленчатого вала более 1100 об./мин. Это обусловлено применением в качестве датчика положения дроссельной заслонки микропереключателя, подающего питание на электропневматический клапан при открытой заслонке. Работа блока управления на холостом ходу идентична блоку 50.3761.
Работа блока управления на холостом ходу идентична блоку 50.3761.
Блок ЭПХХ 1402.3733.
Блок ЭПХХ 1402.3733 устанавливается на автомобили семейства ГАЗ и УАЗ. Принцип его работы такая же как и блока 50,3761. Отличие блоков только в схеме.
Неисправность блока управления ЭПХХ.
При неисправности блока ЭПХХ двигатель не будет работать на холостом ходу или при отпущенной педали газа, обороты скачут от 900 до 1200. Для поиска неисправности достаточно просто удалить сердечник на клапане или соединить трубки на карбюраторе помимо «баллончика».
Система ЭПХХ предназначена для прекращения подачи топлива при следовании автомобиля в режиме принудительного холостого хода. Этот режим характеризуется частотой вращения коленвала двигателя, который превышает обороты холостого хода при закрытой дроссельной заслонке. Такой режим применяется в городском цикле и следовании под гору в режиме торможения двигателем.
На инжекторных двигателях прекращение подачи топлива осуществляет электронная система управления двигателем, а в карбюраторных двигателях блок управления ЭПХХ.
Из чего состоит система ЭПХХ.
В состав система ЭПХХ входит блок управления, электромагнитный клапан или электромагнитный пневмоклапан, датчик положения дроссельной заслонки. В качестве датчика числа оборотов часто используется прерыватель трамблёра.
Датчиком положения дроссельной заслонки может быть микропереключатель, контакты которого размыкаются при закрытой дроссельной заслонке либо датчик-винт на конце, которого крепится провод соединяющий вывод блока управления с массой при открытой дроссельной заслонке.
Датчиком оборотов в карбюраторном двигателе выступает прерыватель трамблёра.
Прекращение подачи топлива осуществляется электромагнитным клапаном или электропневматическим клапаном, в зависимости от конструкции карбюратора. Электромагнитный клапан устанавливается на карбюраторе и перекрывает, при отсутствии питания на нём, своим сердечником канал холостого хода. Электропневматический клапан устанавливается на корпусе автомобиля в разрыве трубки соединяющей впускной коллектор с модулем экономайзера, при включении отключает экономайзер от коллектора и соединяет его с атмосферой.В результате этого клапан экономайзера прекращает подачу топлива.
Принцип работы система ЭПХХ.
При частоте вращения коленвала более 1100 об/мин. и закрытой дроссельной заслонке, блок управления отключает питание с клапана, что прекращает подачу топлива, что позволяет экономить топливо на 2-3% и снизить токсичность выхлопных газов на 15-30%. Кроме этого система ЭПХХ предотвращает детонацию двигателя при отключении двигателя, то есть детонационное сгорание топлива при выключенном зажигании.
Необходимость системы ЭПХХ.
Езда автомобиля с применением принудительного холостого хода происходит крайне редко, в основном в гористой местности. По этому даже обещанные 2-3% практически не достижимая цель. Но предотвращение детонации двигателя это очень часто необходимо. Но для реализации предотвращения детонации при отключении зажигания не обязательно подключения всей схемы. Для этого достаточно просто подать питание на клапан при включении зажигания и снятие его при выключении.
Экономайзер принудительного холостого хода или ЭПХХ позволяет в значительной мере уменьшить выброс токсических веществ в атмосферу. Также он понижает потребление топлива.
Что собой представляет экономайзер
Устройство и схема подключения экономайзера
Устройство ЭПХХ не представляет особой сложности, несмотря на это эффективность системы не поддаётся сомнениям. Стандартная конструкция состоит из таких элементов, как:
- катушка зажигания,
- изолированный наконечник,
- винт,
- электромагнитный клапан,
- блок управления ЭПХХ.
Каждая из этих деталей взаимодействует друг с другом. Результатом подобного процесса является повышенная производительность мотора и существенное увеличение экономии топлива. Но для того чтобы достигнуть такого результата всё должно быть правильно подключено. О том, как это сделать вы можете узнать из схемы подключения ЭПХХ ниже.
Принцип работы
Есть такое понятие, как торможение двигателем. Проще говоря, это ситуация, когда автомобиль продолжает своё движение по инерции. При этом передача всё ещё включена, а педаль, отвечающая за карбюратор отпущена. Подобное состояние также называется принудительным холостым ходом. Отсюда, собственно, и аббревиатура.
При этом внутри двигателя происходят очень интересные и важные процессы. Естественно, топливная смесь в цилиндрах продолжает воспламеняться. Но при этом эффективность работы системы падает в несколько раз. Как результат отработанные газы имеют повышенное содержание оксида углерода и углеводородов.
Внимание! На принудительном холостом ходу топливо расходуется крайне неэкономно.
Естественно, автомобильные инженеры не могли просто так оставить подобный дефект. Результатом долгих исследований и экспериментов стало изобретение системы ЭПХХ. Она позволяет отключить подачу топлива при работе в режиме холостого хода, тем самым решив ряд описанных выше проблем.
Отключение подачи топлива стало возможным благодаря электромагнитному клапану, который монтируется в крышке карбюратора. В данной конструкции за подачу тока отвечает блок управления. Он вместе с клапаном создаёт одну электрическую цепь, в которую также входит:
- источник питания;
- датчик, фиксирующий положение дроссельной заслонки;
- катушка зажигания,
- масса.
Информация передаётся посредством электрического импульса, который идёт от катушки зажигания. Обычно он содержит данные о частоте вращения. О том, что карбюратор перешёл в режим холостого хода сигнализирует датчик. Это третий контакт, который подключается к одному из винтов. Замыкание делается на массу.
Система ЭПХХ работает таким образом, что на холостом ходу обмотка пятого электромагнитного клапана обесточивается. Результатом этого действия является прекращение подачи топлива.
Для того чтобы подача топлива возобновилась система ЭПХХ при помощи второго блока должна зарегистрировать два изменения:
- Частота вращения коленвала должна превысить отметку в 2000 оборотов за минуту.
- Дроссельная заслонка должна находиться в закрытом положении.
Только тогда, когда эти два условия будут выполнены, система ЭПХХ сможет возобновить подачу топлива. Но не всё так просто. Если с пониманием внутренних процессов никаких сложностей возникнуть не должно, то возникает другой закономерный вопрос, а что для этого нужно сделать водителю?
В действительности всё довольно просто. Чтобы система ЭПХХ возобновила подачу топлива водителю нужно совершить некоторые действия. Вначале необходимо уменьшить скорость движения . При этом нельзя нажимать на педаль, которая контролирует положение дроссельной заслонки.
Есть ещё один способ дезактивировать систему ЭПХХ. Для этого вам также нужно вдавить в пол педаль, отвечающую за дроссельную заслонку. Но частота вращения должна быть высокой. Чтобы этого добиться нужно продолжать движение.
Внимание! Система ЭПХХ включает подачу топлива при 150—200 оборотах в минуту.
Отдельно нужно упомянуть особенности работы электромагнитного клапана. Он обесточивается, когда включается зажигание. Подобная предосторожность позволяет исключить то, что двигатель начнёт свою работу с воспламенения.
Неисправности и диагностика ЭПХХ
Система ЭПХХ не отличается особенной сложностью. Именно этот факт выступает гарантией долгой работы. Но даже эта деталь может выйти из строя при больших нагрузках и длительной эксплуатации автомобиля.
Обычно при выходе из строя системы — двигатель не запускается при отпущенной педали. Он просто глохнет. Начать диагностику нужно с проверки шланга, который соединяет пневматический электрический клапан и клапан ЭПХХ.
Внимание! Двигатель может глохнуть из-за того, что в шланге происходит подсос.
Также нужно при диагностике системы ЭПХХ с большим вниманием отнестись к электрическим контактам. Вы должны проверить надёжность соединений. Довольно часто из строя выходит пневматический электрический клапан. Поэтому очень важно осмотреть и его. Следующими на очереди идут ЭБУ и микропереключатель. Проверку можно проводить только при включённом зажигании и неработающем моторе!
Признаком работоспособности пневматического электроклапана будет характерный щелчок, раздающийся при отсоединении и подключении кабеля. Если же этого нет, то дальнейшую проверку нужно осуществлять при помощи контрольной лампы. Это поможет определить есть ли подача тока. При его отсутствии дальше проверяется ЭБУ и микропереключатель.
Итоги
ЭПХХ позволяет добиться значительной экономии топлива. Это крайне выгодное конструктивное решение, позволяющее при минимальных затратах повысить производительность мотора. Отдельным бонусом идёт снижение токсичности отработанных газов.
При движении автомобиля в городе, двигатель около 25 % времени работает на принудительном холостом ходу, когда коленчатый вал двигателя вращается за счет кинетической энергии автомобиля и он движется с включенной передачей и отпущенной педалью управления дроссельной заслонкой. В таких режимах двигатель управляется с помощью экономайзера принудительного холостого хода.
Система управления экономайзера принудительного холостого хода УАЗ, блок управления, клапан, микровыключатель.
На принудительном холостом ходу двигатель расходует топливо, не выполняя полезную работу, в результате быстрого закрытия дроссельной заслонки горючая смесь переобогащается и токсичность отработавших газов увеличивается. Для снижения расхода топлива и токсичности отработавших газов на автомобилях УАЗ установлена электрическая система управления экономайзера принудительного холостого хода (ЭПХХ).
Система управления и электрооборудование экономайзера принудительного холостого хода на автомобилях УАЗ с двигателями УМЗ включает в себя блок управления 1422.3733, электромагнитный клапан 1902.3741 и концевой выключатель карбюратора (микровыключатель) 421.3709.
Принцип работы системы управления экономайзера принудительного холостого хода на автомобилях УАЗ.
Режим принудительного холостого хода характеризуется двумя признаками: частота вращения коленчатого вала двигателя больше частоты холостого хода и дроссельная заслонка карбюратора закрыта. Информация о частоте вращения коленчатого вала двигателя поступает в блок управления ЭПХХ с датчика, в качестве которою используется первичная обмотка катушки зажигания, а информация о закрытии дроссельной заслонки – с концевого выключателя, микровыключателя или датчика-винта .
При отпускании педали акселератора, в следствии переключения контактов концевого выключателя карбюратора блок управления ЭПХХ вырабатывает сигналы управления электромагнитным (электропневматическим) клапаном подачи топлива в зависимости от частоты вращения коленчатого вала двигателя. Если частота вращения коленчатого вала выше частоты холостого хода, то блок управления снимает напряжение с электроклапана, и подача топлива в двигатель прекращается.
При этом частота вращения коленчатого вала снижается, и когда она станет меньше частоты холостого хода, блок управления подключит напряжение бортовой сети к электроклапану. Подача топлива возобновится и частота вращении коленчатого вала возрастет.
При частоте вращения коленчатого вала, снова ставшей больше частоты холостого хода, блок управления опять отключит электроклапан. Процесс повторяется. Периодической отключение подачи топлива в этом режиме снижает расход бензина на 2-3%, а токсичность отработавших газов уменьшается на 15-30%
При нажатии на педаль акселератора контакты концевого выключателя переключаются таким образом, что на электроклапан будет постоянно подаваться напряжение бортовой сети. Топливо при этом будет подаваться независимо от частоты вращения коленчатого вала двигателя.
Блок управления 1422.3733 экономайзера принудительного холостого хода на автомобилях УАЗ, принцип работы.
На автомобилях УАЗ с двигателями УМЗ применяются четырехштырьковые блоки управления 1422.3733 экономайзера. В качестве датчик положения дроссельной заслонки используется микровыключатель 421.3709. При закрытой дроссельной заслонке импульсы напряжения пропорциональные частоте вращения коленчатого вала, поступают с первичной обмотки катушки зажигания 1 на вход полупроводникового ключа, собранного на транзисторе VT1.
Во время действия импульса ключ открывается и конденсатор СЗ разряжается. В паузах между импульсами конденсатор СЗ заряжается. Время заряда, а следовательно, и напряжение на СЗ увеличивается с уменьшением частоты вращения коленчатого вала. При частоте больше частоты холостого хода напряжение на СЗ мало, транзисторы VT2, VT4, VT5, VT6 закрываются. На электромагнитный (электропневматический) клапан напряжение не подается.
Клапан закрывается и подача топлива прекращается. Частота вращения коленчатого вала падает. При частоте меньше частоты холостого хода конденсатор СЗ во время паузы между импульсами успевает зарядиться до напряжения, превышающего опорное напряжение порогового элемента, собранного на транзисторах VT2, VT4. Транзисторы VT2 и VT4 при этом открываются, что обеспечивает открытие транзисторов VT5 и VT6. На электропневмоклапан при этом подается напряжение.
Клапан срабатывает и включает подачу топлива. При открытии дроссельной заслонки контакты микропереключателя S1 замыкаются и напряжение бортовой сети постоянно поступает на электропневмоклапан. Клапан постоянно открыт независимо от сигналов блока управления 1422.3733 экономайзера принудительного холостого хода.
Блок управления ЭПХХ 4-х контактное К-151(25.3761)
Блок управления электроблокировки дверей (замков)Блок управления ЭПХХ 6-ти контактное ДААЗ (УАЗ-4178)Увеличить Внимание! Фотография носит исключительно ознакомительный характер и может отличатся от товара, фактически имеющегося на складе. | Номенклаторный номер: 3741-00-3733000-00 | ||
Позвоните, чтобы уточнить цену
| |||
Для управления клапаном ЭПХХ в карбюраторных двигателях автомобилей используется блок управления ЭПХХ 25.3761. Для включения и отключения электромагнитного клапана экономайзера принудительного холостого хода карбюратора К151. Система ЭПХХ (управления экономайзером принудительного холостого хода) служит для снижения расхода топлива и токсичности отработавших газов. Система состоит из электро-пневмоклапана, блока управления и микровыключателя. Боковой штуцер электропневмоклапана соединен трубкой с пневмоклапаном экономайзера принудительного холостого хода в карбюраторе, а центральный штуцер — с патрубком впускной трубы. Микро выключатель установлен на кронштейне карбюратора, элек-тропневмоклапан и блок управления — на моторном щите. Работает система так. Блок управления непрерывно контролирует частоту вращения коленчатого вала двигателя. При частоте врашения 1500 мин-1±5% и закрытой дроссельной заслонке (режим принудительного холостого хода) блок управления отключает электропневмоклапан, его внутренняя полость сообщается с атмосферой, и связанный с ним пневмоклапан экономайзера карбюратора перекрывает подачу топлива в систему холостого хода. При снижении частоты вращения коленчатого вала двигателя до 1245 мин-1±5% блок управления включает электропневмоклапан, и подача топлива в систему холостого хода возобновляется. При открытой дроссельной заслонке подачу напряжения к электропневмоклапану обеспечивает микровыключатель, на рычажок которого воздействует плечо рычага привода дроссельной заслонки. Неисправность системы ЭПХХ проявляется в рывках автомобиля при движении с малой скоростью, остановках двигателя в режиме холостого хода или вспышках в цилиндрах двигателя после выключения зажигания. | |||
Мнения покупателей:Еще нет мнений об этом товаре.Пожалуйста, войдите, чтобы оставить свое мнение. | |||
Система управления экономайзера принудительного холостого хода
При движении автомобиля в городе, двигатель около 25 % времени работает на принудительном холостом ходу, когда коленчатый вал двигателя вращается за счет кинетической энергии автомобиля и он движется с включенной передачей и отпущенной педалью управления дроссельной заслонкой. В таких режимах двигатель управляется с помощью экономайзера принудительного холостого хода.
Система управления экономайзера принудительного холостого хода УАЗ, блок управления, клапан, микровыключатель.
На принудительном холостом ходу двигатель расходует топливо, не выполняя полезную работу, в результате быстрого закрытия дроссельной заслонки горючая смесь переобогащается и токсичность отработавших газов увеличивается. Для снижения расхода топлива и токсичности отработавших газов на автомобилях УАЗ установлена электрическая система управления экономайзера принудительного холостого хода (ЭПХХ).
Система управления и электрооборудование экономайзера принудительного холостого хода на автомобилях УАЗ с двигателями УМЗ включает в себя блок управления 1422.3733, электромагнитный клапан 1902.3741 и концевой выключатель карбюратора (микровыключатель) 421.3709.
Принцип работы системы управления экономайзера принудительного холостого хода на автомобилях УАЗ.
Режим принудительного холостого хода характеризуется двумя признаками : частота вращения коленчатого вала двигателя больше частоты холостого хода и дроссельная заслонка карбюратора закрыта. Информация о частоте вращения коленчатого вала двигателя поступает в блок управления ЭПХХ с датчика, в качестве которою используется первичная обмотка катушки зажигания, а информация о закрытии дроссельной заслонки — с концевого выключателя, микровыключателя или датчика-винта карбюратора.
Схема соединений электрооборудования экономайзера принудительного холостого хода на автомобилях УАЗ.
При отпускании педали акселератора, в следствии переключения контактов концевого выключателя карбюратора блок управления ЭПХХ вырабатывает сигналы управления электромагнитным (электропневматическим) клапаном подачи топлива в зависимости от частоты вращения коленчатого вала двигателя. Если частота вращения коленчатого вала выше частоты холостого хода, то блок управления снимает напряжение с электроклапана, и подача топлива в двигатель прекращается.
При этом частота вращения коленчатого вала снижается, и когда она станет меньше частоты холостого хода, блок управления подключит напряжение бортовой сети к электроклапану. Подача топлива возобновится и частота вращении коленчатого вала возрастет.
При частоте вращения коленчатого вала, снова ставшей больше частоты холостого хода, блок управления опять отключит электроклапан. Процесс повторяется. Периодической отключение подачи топлива в этом режиме снижает расход бензина на 2-3%, а токсичность отработавших газов уменьшается на 15-30%
При нажатии на педаль акселератора контакты концевого выключателя карбюратора переключаются таким образом, что на электроклапан будет постоянно подаваться напряжение бортовой сети. Топливо при этом будет подаваться независимо от частоты вращения коленчатого вала двигателя.
Блок управления 1422.3733 экономайзера принудительного холостого хода на автомобилях УАЗ, принцип работы.
На автомобилях УАЗ с двигателями УМЗ применяются четырехштырьковые блоки управления 1422.3733 экономайзера. В качестве датчик положения дроссельной заслонки используется микровыключатель 421.3709. При закрытой дроссельной заслонке импульсы напряжения пропорциональные частоте вращения коленчатого вала, поступают с первичной обмотки катушки зажигания 1 на вход полупроводникового ключа, собранного на транзисторе VT1.
Принципиальная схема блока управления 1422.3733 экономайзера принудительного холостого хода на автомобилях УАЗ с двигателями УМЗ.
Во время действия импульса ключ открывается и конденсатор СЗ разряжается. В паузах между импульсами конденсатор СЗ заряжается. Время заряда, а следовательно, и напряжение на СЗ увеличивается с уменьшением частоты вращения коленчатого вала. При частоте больше частоты холостого хода напряжение на СЗ мало, транзисторы VT2, VT4, VT5, VT6 закрываются. На электромагнитный (электропневматический) клапан напряжение не подается.
Клапан закрывается и подача топлива прекращается. Частота вращения коленчатого вала падает. При частоте меньше частоты холостого хода конденсатор СЗ во время паузы между импульсами успевает зарядиться до напряжения, превышающего опорное напряжение порогового элемента, собранного на транзисторах VT2, VT4. Транзисторы VT2 и VT4 при этом открываются, что обеспечивает открытие транзисторов VT5 и VT6. На электропневмоклапан при этом подается напряжение.
Клапан срабатывает и включает подачу топлива. При открытии дроссельной заслонки контакты микропереключателя S1 замыкаются и напряжение бортовой сети постоянно поступает на электропневмоклапан. Клапан постоянно открыт независимо от сигналов блока управления 1422.3733 экономайзера принудительного холостого хода.
Похожие статьи:
- Регуляторы напряжения РР350 и РР132А, характеристики, устройство и принцип действия.
- Генераторы Г250-Е1 и Г250-П2 на УАЗ-469 и УАЗ-3151, характеристики, устройство и принцип действия.
- Датчики уровня жидкостей и снижения давления в контуре тормозов, устройство, принцип действия, проверка исправности.
- Контактная система зажигания УАЗ, состав и общее устройство, схемы контактной системы зажигания.
- Указатель температуры охлаждающей жидкости 14.3807 и датчик ТМ100, проверка и диагностика неисправностей.
- Указатель давления масла 15.3810 и датчик давления ММ358, проверка исправности, основные характеристики.
Система управления экономайзером принудительного холостого хода
Категория:
Автомобили Москвич
Публикация:
Система управления экономайзером принудительного холостого хода
Читать далее:
Система управления экономайзером принудительного холостого хода
Электронная система управления экономайзером принудительного холостого хода, применяемая на автомобилях АЗЛК-2141 и -21412, служит для улучшения экономических параметров работы двигателя и снижения уровня токсичности отработавших газов. Принцип действия ЭПХХ основан на отключении подачи топлива в цилиндры двигателя на режимах принудительного холостого хода, т. е. в тех случаях, когда педаль управления дроссельной заслонкой отпущена, а двигатель остается соединенным с трансмиссией, что влечет за собой повышенную частоту вращения коленчатого вала по сравнению с режимом холостого хода. Этот режим движения автомобиля называется «торможение двигателем», так как от двигателя требуется не отбор мощности, а напротив — он становится дополнительным тормозом. При отсутствии ЭПХХ на этом режиме двигатель продолжает потреблять топливо, причем его расход пропорционален расходу на режиме холостого хода с учетом увеличенной частоты вращения коленчатого вала.
Отключение подачи топлива на режиме принудительного холостого хода приводит к прекращению процесса сгорания смеси в цилиндрах двигателя и торможение двигателем осуществляется более эффективно. Это свойство ЭПХХ особенно ценно при движении автомобиля в условиях горных дорог, где данный прием торможения применяется наиболее часто и результативно.
После включения зажигания система управления ЭПХХ также автоматически отключает подачу топлива, что исключает самовоспламенение смеси в цилиндрах двигателя и неконтролируемую его работу.
Рекламные предложения на основе ваших интересов:
По принципу действия и устройству системы ЭПХХ на обеих моделях автомобилей идентичны и различаются лишь конструктивным исполнением отдельных элементов. Так, для определения положения дроссельной заслонки карбюратора на двигателе автомобиля АЗЛК-2141 применен микропереключатель типа 421.3709, а на двигателе автомобиля АЗЛК-21412 — датчик-винт. Кроме того, на автомобиле АЗЛК-2141 подача топлива отключается с помощью пневмоклапана, расположенного в системе холостого хода карбюратора. На пневмоклапан воздействует электромагнитный клапан, который при наличии на нем напряжения сообщает полость пневмоклапана с впускным трубопроводом, обеспечивая таким образом подачу топлива в цилиндры двигателя. При отсутствии напряжения на обмотке электромагнитного клапана топливо не подается.
На автомобиле АЗЛК-21412 электромагнитный клапан встроен непосредственно в карбюратор и выполнен нормально закрытым, т. е. при протекании тока по обмотке клапана его запорный элемент не препятствует подаче топлива в двигатель, тогда как при обесточенной обмотке клапана его игла перекрывает подачу топлива. Управляется электромагнитный клапан как микропереключателем (датчиком-винтом), установленным на карбюраторе и работающим в зависимости от положения дроссельной заслонки, так и электронным блоком управления ЭПХХ. Напряжение на обмотку электромагнитного клапана от микропереключателя или датчика-винта подается после нажатия на педаль «газа».
Блок управления ЭПХХ подает напряжение на обмотку клапана после падения частоты вращения коленчатого вала ниже определенной величины.
Таким образом, системой холостого хода карбюратора топливо подается либо при низкой частоте вращения коленчатого вала двигателя за счет подачи напряжения на обмотку клапана блоком управления ЭПХХ, либо при нажатии на педаль «газа» за счет микропереключателя или датчика-винта. При отпускании педали «газа» и вращении коленчатого вала с частотой, превышающей 1500 1/мин (для АЗЛК-2141) или 2100 1/мин (для АЗЛК-21412) — порог срабатывания блока управления ЭПХХ — напряжение на клеммах электромагнитного клапана отсутствует и топливо в систему холостого хода карбюратора не подается.
Подача топлива возобновляется при падении частоты вращения коленчатого вала двигателя до 1140 1/мин (АЗЛК-2141) и 1900 1/мин (для АЗЛК-21412) за счет действия блока управления или после нажатия на педаль «газа» — за счет действия микропереключателя или датчика-винта.
В эксплуатации необходимо следить за чистотой элементов системы и надежностью соединений электрических цепей. На практике возможны два вида нарушений работоспособности системы ЭПХХ. Во-первых, при систематической остановке двигателя на режиме холостого хода следует проверить исправность блока управления ЭПХХ или электромагнитного клапана (способы проверки и обнаружения неисправности описаны ниже). Необходимо также убедиться в надежности контакта в соединениях блока управления и электромагнитного клапана. При обнаружении места нарушения контакта выявить причину и устранить неисправность.
Во-вторых, если подача топлива на режимах принудительного холостого хода не прекращается, кроме неисправности блока управления ЭПХХ, возможна разрегулировка датчика-винта или наличие короткого замыкания неподвижного контакта датчика на «массу». Восстановление работоспособности системы производится регулировкой положения датчика-винта или устранением короткого замыкания неподвижного контакта.
Рекламные предложения:
Читать далее: Блок управления ЭПХХ автомобиля «Москвич»
Категория: — Автомобили Москвич
Главная → Справочник → Статьи → Форум
Полиморфизм микросомальной эпоксидгидролазы Tyr113His: связь с риском рака яичников
Функциональное значение было продемонстрировано in vitro для полиморфизма аминокислотной замены экзона 3 T -> C Tyr113His гена микросомальной эпоксидгидролазы (EPHX). Ранее сообщалось, что более высокая активность или быстрый генотип ТТ связаны с повышенным риском рака яичников, и эта ассоциация может отражать усиленную активацию эндогенных или экзогенных субстратов для более реактивных и мутагенных производных.Компоненты сигаретного дыма являются примерами экзогенных субстратов, подверженных такой биоактивации, и воздействие курения может, таким образом, изменять риск, связанный с полиморфизмом EPHX. Мы изучили 545 случаев эпителиального рака яичников и 287 здоровых контрольных случаев этого генетического варианта EPHX T-C, чтобы выяснить, был ли генотип TT в австралийской популяции связан с (i) специфическими характеристиками опухоли яичников; (ii) риск рака яичников в целом или для отдельных подгрупп; и (iii) риск рака яичников, особенно у курильщиков.Генотипирование проводили с использованием системы обнаружения последовательностей Perkin-Elmer ABI Prism 7700 для распознавания аллелей флуорогенной полимеразной цепной реакцией. Стратификация случаев рака яичников в соответствии с поведением опухоли (низкий потенциал злокачественности или инвазивность), степенью, стадией и иммуногистохимическим статусом p53 не продемонстрировала какой-либо гетерогенности в отношении генотипа, определяемого полиморфизмом EPHX. Было высказано предположение о гетерогенности в отношении гистологического подтипа (P = 0,03), в основном из-за снижения частоты генотипа TT в эндометриоидных опухолях.Распределение генотипов EPHX существенно не различается между здоровыми контрольными группами и случаями рака яичников (в целом, с низким потенциалом злокачественности или инвазивным) ни в целом, ни после стратификации по статусу курения. Однако генотип TT был связан со сниженным риском инвазивного рака яичников конкретно эндометриоидного подтипа (отношение шансов с поправкой на возраст = 0,38, 95% доверительный интервал = 0,17-0,87). Результаты показывают, что предложенная EPHX-опосредованная биоактивация компонентов сигаретного дыма до мутагенных форм вряд ли будет участвовать в этиологии рака яичников в целом, но что более высокая скорость EPHX-опосредованной детоксикации может снизить риск эндометриоидного рака яичников.Мол. Канцерогенный. 30: 71-78, 2001.
Полиморфизмы генотипов GSTT1, GSTM1 и EPHX у пациентов с криптогенной полинейропатией: исследование случай – контроль
Brain Behav. 2011 ноя; 1 (2): 135–141.
Йонас Линд
1 Отделение неврологии, Отделение внутренней медицины, Окружная больница Рыхов, Йенчепинг, Швеция
Петер Седерквист
2 Отделение клеточной биологии, Отделение биомедицины и хирургии, Университет Линчёпинга, Линчёпинг Швеция
Матс Фредриксон
3 Кафедра медицины труда и окружающей среды, Университетская больница, Линчёпинг, Швеция
Шахрзад Хоссейниния
4 Кафедра медицины труда и окружающей среды, Медицинский факультет Тегеранского университета, Тегеран, Иран
Мартин Тондель
5 Медицина труда и окружающей среды, Департамент общественного здравоохранения и общественной медицины, Гетеборгский университет, Гетеборг, Швеция
Бодил Перссон
3 Департамент медицины труда и окружающей среды, Университетская клиника, Линчёпинг, Швеция
Магну s Vrethem
6 Отделения неврологии и нейрофизиологии, Университетская клиника Линчёпинга, Линчёпинг, Швеция
1 Отделение неврологии, Отделение внутренней медицины, Окружная больница Рихов, Йёнчёпинг, Швеция
2 Отдел клеточной Биология, кафедра биомедицины и хирургии, Университет Линчёпинга, Линчёпинг, Швеция
3 Кафедра медицины труда и окружающей среды, Университетская больница, Линчёпинг, Швеция
4 Кафедра медицины труда и окружающей среды, Медицинский факультет Университета Тегеран, Тегеран, Иран
5 Медицина труда и окружающей среды, Департамент общественного здравоохранения и общественной медицины, Гетеборгский университет, Гетеборг, Швеция
6 Кафедры неврологии и нейрофизиологии, Университетская больница Линчёпинга, Линчёпинг, Швеция
Джонас Линд, Отделение неврологии, Депутат т внутренних болезней, окружная больница Рыхов, S-551 85 Йёнчёпинг, Швеция.Тел: +4636322020; Факс: +4636322188; Электронная почта: [email protected].Финансируется за счет грантов FORSS (Совет медицинских исследований Юго-Восточной Швеции) и Futurum — академии здравоохранения при совете графства Йёнчёпинг.
Поступило 14.06.2011; Пересмотрено 2 сентября 2011 г .; Принято 12 сентября 2011 г.
Copyright © 2011 Авторы. Опубликовано Wiley Periodicals, Inc. Эта статья цитировалась в других статьях в PMC.Abstract
Целью данного исследования было проанализировать, могут ли полиморфизмы нулевых аллелей глутатион-S-трансферазы Mu-1 (GSTM1), глутатион-S-трансферазы тета-1 (GSTT1) и генетической вариации эпоксида с низкой активностью экзон три гидролазы (EPHX * 3) влияет на риск развития полинейропатии.Ферменты этих генов играют важную роль в метаболизме токсичных соединений. Семьдесят девять пациентов с криптогенной полинейропатией (эквивалентной хронической идиопатической аксональной нейропатии) и 398 контрольных пациентов были протестированы на генетический полиморфизм. Медицинские карты были изучены для сбора данных о клинических данных при постановке диагноза, а данные о воздействии были собраны с помощью анкет. Отношения шансов (OR) для нулевых форм GSTM1 и GSTT1 и YY-формы нормальной активности EPHX * 3 были близки к единице, за исключением GSTT1, который достиг 1.86. Самый высокий риск полинейропатии был обнаружен у курильщиков с нулевым GSTT1, у которых риск повышался в 3,7 раза. Взаимодействия между генами были проанализированы и подтвердили увеличение OR для GSTT1, которое было наиболее сильным, если пациенты имели низкоактивную форму HH EPHX * 3 (OR 2,37). Наша гипотеза состоит в том, что нулевой полиморфизм GSTT1 может быть связан с нарушением метаболизма токсичных веществ, что может привести к повреждению нервов в периферической нервной системе.
Ключевые слова: Хроническая идиопатическая аксональная нейропатия, глутатион-S-трансфераза, эпоксидгидролаза, курение, растворители
Введение
Полинейропатия — это неврологическое заболевание, которое часто встречается в среднем и позднем взрослом возрасте.Оценки его распространенности варьируются от 2,4% до 8% и зависят от выбора пациентов, то есть от общей популяции или серии больниц, и даже выше у лиц, подвергшихся воздействию различных токсинов, или пациентов с сахарным диабетом (Beghi et al. 1995 ). Хотя существует несколько известных причин полинейропатии, этиология часто остается неизвестной (Martyn and Hughes, 1997), и тогда ее называют криптогенной. Механизмом в этих случаях часто считается воздействие профессиональных факторов или факторов окружающей среды, которые мы исследовали ранее (Tondel et al.2006 г.). Аксонопатия — наиболее частая форма патологии при токсических невропатиях, но основные патологические механизмы неясны. Считается, что нарушение аксонального транспорта приводит к дегенерации уязвимых дистальных нервных сегментов (Spencer et al. 1979; Griffin and Watson 1988), и когда процесс продолжается, дегенерация продолжается проксимально по направлению к телу клетки.
В промышленных условиях и в окружающей среде некоторые соединения нейротоксичны и часто поражают периферические нервы (Spencer and Schaumburg 2000).Одним из примеров является н-гексан, который накапливается в нервной ткани во время хронического воздействия (Feldman 1999). Известно, что он вызывает первичную дегенерацию аксонов при вторичной демиелинизации (Chang et al. 1993).
Биотрансформация экзогенных и эндогенных соединений может играть роль в индивидуальной восприимчивости из-за генетической изменчивости ферментов, участвующих в детоксикации. Метаболизм биотрансформации можно разделить на две фазы. В фазе 1 исходная чужеродная молекула изменяется путем добавления функциональной группы, которая затем может быть сопряжена в фазе 2.Затем конъюгированная молекула может выводиться из организма (Timbrell 2000). Обычно эти шаги приводят к менее токсичной молекуле, но в некоторых случаях происходит обратное.
Эпоксидгидролазы (EPHX) представляют собой пример ферментной системы фазы 1, которая действует путем добавления воды к эпоксиду (Timbrell 2000). Эти ферменты играют важную роль в метаболизме экзогенных химических веществ, таких как полициклические ароматические углеводороды (ПАУ) (Omiecinski et al. 1993). Эпоксиды могут быть частично детоксифицированы микросомальным EPHX (mEPHX), который катализирует их гидролиз с образованием соответствующих дигидродиолов (Oesch 1973).Хотя этот гидролиз обычно считается реакцией детоксикации, поскольку образуются менее токсичные химические вещества, некоторые дигидродиолы, образующиеся из ПАУ, являются субстратами для дополнительных метаболических изменений высокотоксичных, мутагенных и канцерогенных эпоксидов полициклических углеводородных диолов. Таким образом, EPHX * 3 играет ту же двойную роль в детоксикации и активации проканцерогенов, что и в некоторых цитохромах P450 (Benhamou et al. 1998), и, как следствие, может также играть важную роль в нейротоксичности (Guengerich 1982) и в лекарственном препарате. -связанные побочные эффекты.Два аминокислотных полиморфизма были идентифицированы в кодирующей области экзона три (EPHX * 3), обмена тирозина 113 гистидина (Y113H), что приводит к низкой активности формы фермента (Hassett et al. 1994), которая может влиять на эпоксид. дезактивация в клетке. Пациенты с наследственной оптической невропатией Лебера, гомозиготные по гистидину 113, заболевали раньше, чем пациенты без этого генотипа (Ishikawa et al. 2005). Полиморфизм в экзоне четыре, гистидин 139 аргинин (h239R, rs2234922), был предложен как высокоактивная изоформа mEPHX (Smith and Harrison 1997; Benhamou et al.1998).
Глутатион S-трансферазы (GST) представляют собой семейство ферментов фазы 2, ответственных за метаболизм широкого спектра ксенобиотиков и канцерогенов (Mannervik 1985). Эти ферменты катализируют конъюгацию глутатиона с широким спектром органических соединений с образованием тиоэфиров, реакцию, которая иногда является стадией процесса детоксикации, приводящей к образованию меркаптуровой кислоты, классического продукта экскреции ксенобиотиков (Mannervik 1985). Было показано, что ферменты GST защищают организмы от повреждения реактивными кислородными соединениями благодаря своей способности связываться с глутатионом (Hayes and Strange 2000).На основании сходства аминокислотных последовательностей и перекрестной реактивности антител GST делятся на несколько классов, включая мю и тета. Глутатион-S-трансфераза Mu-1 (GSTM1) и глутатион-S-трансфераза Theta-1 (GSTT1) являются полиморфными у человека, и делеции в генах приводят к фактическому отсутствию активности фермента, особенно с одновременными делециями в генах GSTM1 и GSTT1 ( Абу-Амеро и др., 2009). Генетические вариации могут изменить восприимчивость человека к канцерогенным веществам и токсинам, а также повлиять на токсичность и эффективность некоторых лекарств (Ginsberg et al.2009 г.). Гены, кодирующие класс ферментов mu, как известно, очень полиморфны (Xu et al. 1998). Они участвуют в детоксикации электрофильных соединений, включая канцерогены, терапевтические препараты, токсины окружающей среды и продукты окислительного стресса, путем конъюгации с глутатионом. Сообщалось, что люди с нулевым генотипом GSTM1 и высоким воздействием растворителей подвергаются повышенному риску развития хронической токсической энцефалопатии, вызванной растворителем (Söderkvist et al. 1996) и болезни Паркинсона (Dick et al.2007). Ген GSTT1 расположен на хромосоме 22. Как для GSTT1, так и для GSTM1 нулевой генотип был связан с повышенным риском оптических невропатий (Abu-Amero et al. 2009) и побочных эффектов лекарств, включая когнитивные нарушения после терапии у пациентов. с медуллобластомой (Barahmani et al. 2009), но не с наследственной оптической невропатией Лебера (Ishikawa et al. 2005) или невропатией у пациентов, получающих химиотерапию на основе оксалиплатина (Lecomte et al. 2006).
Поскольку активность этих ферментов, метаболизирующих ксенобиотики, как правило, необходима для обеспечения эффективной детоксикации, тем самым защищая организм от повреждений, вызванных воздействием, мы проанализировали наличие полиморфизма нулевых аллелей GSTM1 и GSTT1 и генетической вариации mEPHX (низкая активность) влияют на риск развития полинейропатии.Материал и методы Больница, Йёнчёпинг) в возрасте от 40 до 79 лет на момент постановки диагноза (Lindh et al. 2005). Было получено одобрение комитета по этике для повторной проверки записей и контакта с этими субъектами.Медицинские карты были повторно изучены с использованием заранее определенного протокола исследования, включая симптомы, признаки и лабораторные тесты, для подтверждения правильного диагноза в каждом случае. Пациенты с доминирующей демиелинизирующей нейропатией, наследственной нейропатией или любой другой выявленной причиной нейропатии были исключены (Lindh et al. 2005).
Из первоначальной когорты из 168 пациентов 158 были еще живы, и их попросили принять участие в исследовании. Образцы крови были взяты у 79 пациентов (ответ 50%), которые согласились участвовать.Было 54 мужчины и 25 женщин с полинейропатией (средний возраст 71,0 и 68,5 лет соответственно). 89 пациентов, которые не участвовали, были немного старше (72,5 против 70,2 года), имели более высокую клиническую (1,6 против 1,4) и нейрофизиологическую тяжесть (2,0 против 1,8), но различия не достигли статистической значимости, и наблюдались нет разницы в половом распределении.
Контрольную группу составили 398 человек из популяционной контрольной группы из шведской части исследования болезни Паркинсона тех же генетических полиморфизмов, живущих в той же географической области (Dick et al.2007). Контрольную группу составили 198 мужчин и 200 женщин (средний возраст 67,4 и 67,5 лет соответственно). Средний возраст среди случаев был немного выше, чем в контроле (70,2 против 67,5 лет, P <0,05).
Заполненные анкеты для пациентов и контрольных групп о статусе курения и других воздействиях, включая воздействие растворителей или пестицидов, общую анестезию и питьевую воду из частных колодцев, как описано ранее (Tondel et al. 2006; Dick et al. 2007).
Клиническая тяжесть неврологического состояния оценивалась на основе функционального дефицита.Клиническая тяжесть 1 степени (легкая) определялась как незначительные двигательные и / или сенсорные симптомы без функционального дефицита. Степень 3 была определена как тяжелые симптомы с функциональным дефицитом, включая легкую атаксию или, по крайней мере, некоторую потребность в помощи. Степень 2 или умеренная степень тяжести определялась как те симптомы и нарушения, которые находились между 1 и 3 степенью. Точно так же пациенты считались имеющими нейрофизиологические данные 1 степени (легкие), если нейрография и ЭМГ (электромиография) при постановке диагноза показывали незначительное снижение сложных моторных аксональных потенциалов (CMAP), аксональных потенциалов сенсорных нервов (SNAP) или скорости проводимости (CV) как минимум в двух нервах.Нейрофизиологические результаты 3 степени (тяжелые) были определены как потеря сенсорных или моторных реакций по крайней мере в двух нервах, как оценивалось в предыдущем исследовании, а степень 2 (умеренная) — как нейрофизиологические результаты между 1 и 3 уровнями (Lindh et al. 2005). .
Собирали цельную кровь и выделяли лейкоцитарную ДНК с помощью набора для очистки ДНК Wizard Genome (Promega Inc., Мэдисон, Висконсин). Нулевые генотипы GSTM1 и GSTT1 оценивали с помощью мультиплексной полимеразной цепной реакции (ПЦР) с β-глобином в качестве внутреннего контрольного гена для успешной амплификации ПЦР (Arand et al.1996). Полиморфизм аминокислот в гене mEPHX (экзон 3 EPHX1) определяли с помощью анализа PCR-RFLP (полиморфизм длины рестрикционного фрагмента) (Lancaster et al. 1996; Smith and Harrison 1997). Для экзона 3 существует три возможных генотипа: YY, YH и HH. Аллель нормальной активности дикого типа — YY, а генотип низкой активности — HH.
Комитет по этике факультета медицинских наук Университета Линчёпинга одобрил проект.
Статистические методы
Статистический анализ проводился с использованием SPSS версии 15.Поскольку ни контрольная группа, ни пациенты с полинейропатией не были нормально распределены по возрасту, статистический анализ проводился с использованием непараметрического метода; тест Крускала – Уоллиса с последующим U-критерием Манна – Уитни для апостериорного анализа (с использованием поправки Бонферони для множественных анализов). Для категориальных переменных использовался критерий хи-квадрат. Для групп, состоящих менее чем из пяти респондентов, анализ проводился с использованием точного критерия Фишера. Относительный риск выражали как отношение шансов (ОШ) с 95% доверительным интервалом (ДИ).Сравнения считались значимыми, если значения P были <0,05. Полиморфизм анализировали независимо от пола, поскольку гены расположены на аутосомах.
Анализ множественной логистической регрессии использовался для получения оценок OR для генетических факторов с поправкой на следующие заранее заданные ковариаты: возраст, пол, когда-либо употреблявшиеся табак, воздействие растворителей или пестицидов, общая анестезия и питьевая вода из частных колодцев. Взаимодействие между генами анализировали с помощью Stata версии 11.0.
Результаты
Всего 79 случаев с криптогенной полинейропатией и 398 контрольных пациентов были протестированы на генетический полиморфизм в генах GSTM1, GSTT1 и mEPHX. Частоты различных генетических полиморфизмов представлены в. Среди контрольной группы было значительно больше людей с нулевым GSTT1 у женщин, чем у мужчин ( P = 0,04), а гомозиготный вариант HH в mEPHX чаще встречался у мужчин ( P <0,01). Остальные варианты у мужчин и женщин не различались.Статистически значимых различий между случаями и контролем не было ни в одной группе.
Таблица 1
Распределение генетических полиморфизмов у пациентов с криптогенной полинейропатией и контрольной группы
| Общее количество испытуемых | Мужчины | Женщины | |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 9014 9014 9014 9014 9014 9014 9014 9014 9014 9014 9014 | Pnp | Элементы управления | Pnp | Элементы управления | Pnp | Элементы управления | |||||||||||||||||||||||||||||||
| 9014 9014 9014 9014 9014 9014 9014 9014 9016 9014 9014 9014 9014 9016 | % | n | % | n | % | n | % | n | % | % | 40 | 50.6 | 201 | 50,5 | 30 | 55,6 | 100 | 50,5 | 10 | 40,0 | 101 | 50,5 | |||||||||||||||
| GSTM4 | 24 | 44,4 | 98 | 49,5 | 15 | 60,0 | 99 | 49,5 | |||||||||||||||||||||||||||||
| GSTT1 null | 7 | 8,9 | 613 | 3 | 5,6 | 23 | 11,6 | 4 | 16,0 | 38 | 19,0 | ||||||||||||||||||||||||||
| GSTT1 pos | 72 | 91,1 9024 5 | 175 | 88,4 | 21 | 84,0 | 162 | 81,0 | |||||||||||||||||||||||||||||
| EPHX * 3 YH | 30 | 38,0 | 142 | 35,7 | 369 | 34,8 | 12 | 48,0 | 73 | 36,5 | |||||||||||||||||||||||||||
| EPHX * 3 YY | 39 | 49,4 | 202 | 9024 9024 47,424 | 16,0 | 18 | 9,0 | ||||||||||||||||||||||||||||||
OR для нулевых форм GSTM1 и GSTT1 и YY-формы EPHX * 3 были близки к единице для всех полиморфизмов, кроме GSTT1, которая достигла 1,86. Когда мужчин и женщин анализировали отдельно, мы обнаружили, что OR EPHX * 3 YH и HH по сравнению с YY составляло 0,7 у мужчин, тогда как у женщин оно составляло 2,1, почти достигая значимости ().
Таблица 2
Анализ генетических факторов по статусу случай-контроль (случаи vs.контрольные)
| Все испытуемые | Мужчины | Женщины | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 9014 9014 9014 9014 Исследуемые группы | OR | P Значение | OR | P Значение | OR | P Значение | ||||||||
| GSTM1 | Null99 (0,61–1,61) | 1 | 0,82 (0,45–1,50) | 0,54 | 1,53 (0,66–3,57) | 0,40 | ||||||||
| GSTT1 | Null | положительный 1,124 | 0,16 | 2,23 (0,64–7,74) | 0,31 | 1,23 (0,40–3,80) | 1 | |||||||
| EPHX * 3 | YY | YH | 0,64–7,74,09 (1,09)0,81 (0,42–1,57) | 0,62 | 1.99 (0,80–4,50) | 0,16 | ||||||||
| YY | HH | 0,96 (0,45–2,05) | 1 | 0,52 (0,20–1,35) | 0,20 | 0,12 | ||||||||
| YY | YH / HH | 1,06 (0,65–1,71) | 0,90 | 0,71 (0,39–1,30) | 0,29 | 2,13 9024–5 9024–5 | ||||||||
Что касается клинических данных, то у 24 пациентов были признаны умеренные симптомы, а у 39 пациентов — тяжелые.Не было обнаружено значительных различий между группами по клинической или нейрофизиологической тяжести на момент постановки диагноза, за исключением тенденции для нулевого GSTM1 иметь более серьезные клинические проявления, чем для положительных случаев GSTM1 (среднее значение 1,55 против 1,31, P = 0,064). Аксональная нейропатия наблюдалась у 41 пациента, а комбинированная аксональная и демиелинизирующая нейропатия — у 19 пациентов. Что касается нейрофизиологических данных, у двух пациентов была чистая моторная нейропатия, у 13 пациентов была чисто сенсорная нейропатия и у 64 пациентов была смешанная сенсомоторная нейропатия.Генетические полиморфизмы не были существенно связаны с этими нейрографическими находками.
Мы также исследовали эффекты различных воздействий. В контрольной группе было 189 (47%) курильщиков или бывших курильщиков по сравнению с 43 (54%) курильщиками среди пациентов с криптогенной полинейропатией. О воздействии растворителей во время работы или в свободное время сообщили 24 (30%) пациентов с криптогенной полинейропатией и 132 (33%) пациентов из контрольной группы. О воздействии пестицидов сообщили восемь (10%) пациентов с криптогенной полинейропатией и 29 (7%) пациентов из контрольной группы.Всего 59 (74%) пациентов с криптогенной полинейропатией и 312 (78%) пациентов контрольной группы подверглись общей анестезии, 51 (65%) пациентов с криптогенной полинейропатией и 29 (71%) пациентов с криптогенной полинейропатией. органы управления имели частное водоснабжение, по крайней мере, в течение определенного периода своей жизни. OR для криптогенной полинейропатии среди лиц, подвергшихся воздействию, показаны на. GSTT1 null среди курильщиков достиг наивысшего OR (3,72, P = 0,08), а EPHX * 3 HH по сравнению с YY среди подвергшихся воздействию растворителя имел самый низкий OR (0.30, P = 0,14). Логистический регрессионный анализ для различных полиморфизмов, пола, возраста и воздействия не выявил каких-либо искажающих эффектов, за исключением того, что увеличение возраста и мужского пола увеличивало риск криптогенной полинейропатии. Взаимодействия между генами были проанализированы и подтвердили увеличение OR для GSTT1, которое было самым сильным, если у пациентов была HH-форма EPHX * 3 (OR 2.37).
Таблица 3
Эффекты генетических полиморфизмов при различных воздействиях (подверженные случаи и контрольные)
| Генетические полиморфизмы | Исследуемые группы | Воздействие | Отношение шансов | P Величина| Нулевое против положительного | Курение | 1.08 (0,56–2,10) | 0,87 | |
|---|---|---|---|---|---|---|---|---|
| GSTT1 | Нулевое против положительного | Курение | 3,72 (0,85–16,2) | 0,08 | ||||
| EPHX * 3 | Y 1,40 (0,69–2,82) | 0,38 | ||||||
| HH по сравнению с YY | Курение | 0,57 (0,18–1,82) | 0,44 | |||||
| YH / HH по сравнению с 114 (0,58–2,22) | 0,74 | |||||||
| GSTM1 | Нулевой по сравнению с положительным | Растворитель | 0,46 (0,18–1,14) | 0,12 | ||||
| GSTT1 | по положительному 0,28–6,1) | 1 | ||||||
| EPHX * 3 | YH по сравнению с YY | Растворитель | 0,72 (0,27–1,91) | 0,63 | ||||
| 9024 H24 H по сравнению с | 30 (0,06–1,40)0,14 | |||||||
| YH / HH в сравнении с YY | Растворитель | 0,55 (0,22–1,34) | 0,27 | |||||
| GSTM424 Null | положительный (0,19–4,94)1 | |||||||
| GSTT1 | Нулевой по сравнению с положительным | Пестициды | 0,25 (0,01–4,51) | 0,39 | ||||
| EPHX4 9024H1 9024H | .33 (0,24–7,28)1 | |||||||
| HH по сравнению с YY | Пестициды | 0,80 (0,07–9,67) | 1 | |||||
| YH244 | 0,23–5,89) | 1 | ||||||
| GSTM1 | Нулевой по сравнению с положительным результатом | Общая анестезия | 1,09 (0,62–1,90) | 0,78 | ||||
| GSTT4 1 | 9024 по сравнению с общим положительным результатом | 05 (0,94–1,17)0,55 | ||||||
| EPHX * 3 | YH по сравнению с YY | Общая анестезия | 1,10 (0,60–2,11) | 0,76 | ||||
| H245 | Общая анестезия0,95 (0,39–2,32) | 1 | ||||||
| YH / HH по сравнению с YY | Общая анестезия | 1,06 (0,61–1,85) | 0,89 | |||||
| GST по сравнению с положительным | 0.78 (0,43–1,43) | 0,45 | ||||||
| GSTT1 | Нуль по сравнению с положительным значением | Частная вода | 1,66 (0,57–4,91) | 0,48 | ||||
| 1,11 (0,58–2,14) | 1 | |||||||
| HH по сравнению с YY | Частная вода | 1,01 (0,41–2,50) | 1 | |||||
| HH | YH по сравнению с YH | 1.09 (0,60–1,97) | 0,88 |
Обсуждение
В этом эпидемиологическом исследовании случай – контроль пациентов с криптогенной полинейропатией мы изучили связь нулевых полиморфизмов GSTM1 и GSTT1 и нескольких полиморфизмов HH экзона 3 EPHX1. экологическое и химическое воздействие. Хотя мы не обнаружили статистически значимого увеличения риска, нулевой генотип GSTT1 был связан с почти двукратным повышением риска полинейропатии.Наша гипотеза заключается в том, что нулевой полиморфизм GSTT1 может быть связан с нарушением метаболизма токсичных веществ и реактивного кислорода, что может привести к повреждению нервов, затрагивая несколько участков вдоль моторных и сенсорных аксонов в периферической нервной системе. Это может привести к атрофии аксонов или набуханию аксонов, что приведет к прогрессирующей дистальной дегенерации аксонов. Миелиновая оболочка может разрушаться одновременно с аксоном. Это может способствовать или прямо приводить к аксональной или комбинированной аксонально-демиелинизирующей нейропатии.
Компоненты сигаретного дыма являются примерами экзогенных субстратов, которые токсичны и, кроме того, подвержены биоактивации и могут как прямо, так и косвенно быть нейротоксичными. Мы обнаружили почти четырехкратное повышение риска полинейропатии у курильщиков без GSTT1, что почти достигло статистической значимости. Teunissen и соавторы сообщили о OR 2,1 для текущего курения у пациентов с хронической идиопатической аксональной полинейропатией (Teunissen et al., 2002), а также было обнаружено, что употребление табака может предрасполагать к более раннему развитию и более серьезным симптомам диабетической невропатии ( Tesfaye et al.2005). Наши данные показывают, что этот риск может быть объяснен курильщиками, несущими определенные генетические полиморфизмы, приводящие к нарушению детоксикации токсичных соединений в сигаретном дыме. В исследовании хронической токсической энцефалопатии, вызванной растворителем, у курильщиков был обнаружен повышенный коэффициент риска 2,5 для нулевого генотипа GSTM1 и 1,5 для нулевого генотипа GSTT1 в общей популяции. У некурящих нулевой генотип GSTM1 не представлял риска хронической токсической энцефалопатии.Ни один из изученных полиморфизмов mEPHX не был связан с повышенным риском. Механизм токсического воздействия сигаретного дыма на нервную систему не известен, но предполагалось, что он опосредован химическими веществами в дыме, наиболее важным компонентом которого считаются ПАУ. Нарушение распада ПАУ у людей с нулевым генотипом GST может привести к усилению воздействия на нервы и, таким образом, к повышенной потере нервов. н-гексан — еще одно токсичное вещество, которое присутствует в сигаретном дыме и, как хорошо известно, вызывает полинейропатию (Zhang et al.2006 г.). Однако в нашем исследовании курение не увеличивало риск полинейропатии. Усиленная отрицательная связь между курением сигарет и активностью GSTM1 была описана при болезни Паркинсона, которая может быть опосредована нейропротективным действием никотина на дофаминергическую систему (De Palma et al. 2010).
Частота нулевого GSTM1 составляет около 42–60% у кавказцев (Garte et al. 2001). Частота гомозиготного нулевого GSTT1 сильно варьируется в зависимости от этнической принадлежности и составляет 10–20% у европеоидов (Rebbeck 1997).Ген EPHX * 3 можно найти в трех различных формах: генотипы дикого типа / с нормальной активностью (YY), гетерозиготные (YH) или гомозиготные / с низкой активностью (HH). В европеоидной популяции около 40% субъектов являются гетерозиготными, а 12% — гомозиготными по генотипу HH (Garte et al. 2001). Частота этих полиморфизмов в нашей исследуемой популяции была аналогичной. Никаких различий в частотах аллелей по возрасту или полу в крупных исследованиях не наблюдалось (Garte et al. 2001). К сожалению, у нас был дисбаланс в нашей контрольной группе: 19% женщин и 12% мужчин имели нулевой полиморфизм GSTT1, а 9% женщин и 18% мужчин имели полиморфизм EPHX * 3 HH.Таким образом, невозможно сделать какие-либо выводы о различиях в рисках криптогенной полинейропатии у мужчин и женщин по отдельности.
Лица, несущие гены, кодирующие белки с утраченной или нарушенной функцией, имеют нарушенную метаболическую способность выводить токсичные соединения и, следовательно, могут подвергаться повышенному риску полинейропатии. Этот тип мутации часто имеет OR 2–3 для повышенного риска рака. В нашей группе пациентов с полинейропатией мы обнаружили тенденцию к более низкому OR для гена EPHX * 3 по сравнению с контролем.Общий риск полинейропатии, вероятно, является результатом сложного взаимодействия множества генетических факторов и факторов окружающей среды с течением времени. Ранее мы подтвердили, что производственное воздействие растворителя Стоддарда, выхлопных газов бензина, гербицидов или вибраций рук и ног значительно увеличило риск полинейропатии, а также были выявлены новые детерминанты, то есть диоксид серы, ксилол и метилэтилкетон (Tondel et al. 2006 г.).
Мы не обнаружили какой-либо существенной корреляции между клинической или нейрофизиологической серьезностью и генотипом, за исключением небольшого увеличения тяжести клинических проявлений у пациентов с нулевым GSTM1, которые почти достигли статистической значимости.Возможно, что корреляция может быть обнаружена, если будет использована более чувствительная шкала клинической или нейрофизиологической степени тяжести.
Возможной причиной того, что мы не обнаружили существенных различий, является небольшое количество пациентов. В китайском исследовании 22 случаев полинейропатии на полиграфической фабрике и 163 контрольных случаях была обнаружена связь с CYP2E1 Dra, но не с GSTM1 и GSTT1, что указывает на то, что необходимо более выраженное воздействие токсичного агента или что другие гены могут имеют большее значение для развития полинейропатии (Zhang et al.2006 г.). В нашу выборку вошли пациенты с диагнозом криптогенная полинейропатия в отделениях неврологии. Вполне вероятно, что врачи общей практики правильно диагностировали людей, работающих в промышленных условиях с высоким уровнем воздействия токсичных веществ, или что невролог диагностировал у них токсическую невропатию, что привело к недооценке риска воздействия в нашем исследовании.
Решением этих проблем могло бы стать проведение Общегеномного исследования ассоциации (GWAS), которое стало успешным способом поиска новых генов-кандидатов, например, при болезни Паркинсона, болезни Альцгеймера (Gandhi and Wood 2010) и спорадический боковой амиотрофический склероз (Шатунов и др.2010). Однако для этого потребуется очень большое количество пациентов из нескольких стран.
В заключение, не было обнаружено значительной корреляции между полиморфизмами GSTM1, GSTT1 и EPHX1 у пациентов с криптогенной полинейропатией по сравнению с контрольной группой. Однако сильная тенденция наблюдалась в отношении нулевого фенотипа GSTT1 и курения у этих пациентов по сравнению с контрольной группой (OR 3,7). Нулевой полиморфизм GSTT1 может быть связан с нарушением метаболизма токсичных веществ и реактивного кислорода, что может привести к повреждению нервов в периферической нервной системе.Это может способствовать или прямо приводить к аксональной или комбинированной аксонально-демиелинизирующей нейропатии.
Благодарности
Это исследование было поддержано грантами FORSS (Совет медицинских исследований Юго-Восточной Швеции) и Futurum — академии здравоохранения при совете графства Йёнчёпинг.
Ссылки
- Abu-Amero KK, Milcarek B, Bosley TM. Генотипы с делецией GSTM1 и GSTT1 при различных спонтанных оптических невропатиях у арабов. Br. J. Ophthalmol. 2009; 93: 1101–1104.[PubMed] [Google Scholar]
- Arand M, Muhlbauer R, Hengstler J, Jager E, Fuchs J, Winkler L, Oesch F. Протокол мультиплексной полимеразной цепной реакции для одновременного анализа полиморфизмов GSTM1 и GSTT1 глутатион-S-трансферазы . Анальный. Biochem. 1996. 236: 184–186. [PubMed] [Google Scholar]
- Barahmani N, Carpentieri S, Li XN, Wang T, Cao Y, Howe L, Kilburn L, Chintagumpala M, Lau C, Okcu MF. Полиморфизм глутатион-S-трансферазы M1 и T1 может предсказывать побочные эффекты после терапии у детей с медуллобластомой.Neuro. Онкол. 2009; 11: 292–300. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Beghi E, Monticelli ML, (IGPSG) IGPSG Хроническая симметричная симптоматическая полинейропатия у пожилых людей: полевое скрининговое исследование в двух регионах Италии. I. Распространенность и общие характеристики выборки. Неврология. 1995; 45: 1832–1836. [PubMed] [Google Scholar]
- Бенхаму С., Рейникайнен М., Бушарди С., Дайер П., Хирвонен А. Связь между раком легких и генотипами микросомальной эпоксидгидролазы.Cancer Res. 1998. 58: 5291–5293. [PubMed] [Google Scholar]
- Чанг CM, Yu CW, Fong KY, Leung SY, Tsin TW, Yu YL, Cheung TF, Chan SY. Н-гексановая нейропатия в офсетных принтерах. J. Neurol. Нейрохирургия. Психиатрия. 1993; 56: 538–542. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Де Пальма Дж., Дик Ф. Д., Кальцетти С., Скотт Н. В., Прескотт Дж. Дж., Осборн А., Хайтес Н., Моззони П., Негротти А., Скаглиони А. и др. Исследование болезни Паркинсона и употребления табака методом случай-контроль: взаимодействие генов и табака. Mov.Disord. 2010; 25: 912–919. [PubMed] [Google Scholar]
- Дик Ф.Д., Де Пальма Дж., Ахмади А., Осборн А., Скотт Н. В., Прескотт Дж. Дж., Беннет Дж., Семпл С., Дик С., Моццони П. и др. Взаимодействие генов и окружающей среды при паркинсонизме и болезни Паркинсона: исследование Geoparkinson. Ок. Environ. Med. 2007. 64: 673–680. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Feldman RG. н-гексан и метил-н-бутилкетон. В кн .: Профессиональная нейротоксикология и нейротоксикология окружающей среды. Филадельфия, Пенсильвания: Липпинкотт-Рэйвен; 1999 г.С. 290–317. [Google Scholar]
- Gandhi S, Wood NW. Полногеномные исследования ассоциации: ключ к разгадке нейродегенерации? Nat. Neurosci. 2010. 13: 789–794. [PubMed] [Google Scholar]
- Гарте С., Гаспари Л., Александри А. К., Амброзон С., Отруп Х., Отруп Дж. Л., Баранова Х., Батум Л., Бенхаму С., Боффетта П. и др. Частота метаболического полиморфизма генов в контрольных популяциях. Cancer Epidemiol. Биомаркеры Пред. 2001; 10: 1239–1248. [PubMed] [Google Scholar]
- Ginsberg G, Smolenski S, Hattis D, Guyton KZ, Johns DO, Sonawane B.Генетический полиморфизм глутатионтрансфераз (GST): популяционное распределение активности конъюгирования GSTM1, T1 и P1. J. Toxicol. Environ. Здоровье B: крит. Ред. 2009; 12: 389–439. [PubMed] [Google Scholar]
- Гриффин Дж. У., Уотсон Д. Ф. Аксональный транспорт при неврологических заболеваниях. Аня. Neurol. 1988; 23: 3–13. [PubMed] [Google Scholar]
- Guengerich FP. Эпоксидгидролаза: свойства и метаболическая роль. Rev. Biochem. Toxicol. 1982; 4: 4–30. [Google Scholar]
- Hassett C, Aicher L, Sidhu JS, Omiecinski CJ.Человеческая микросомальная эпоксидгидролаза: генетический полиморфизм и функциональная экспрессия in vitro вариантов аминокислот. [Ошибка появляется в Hum Mol Genet 1994 Jul; 3 (7): 1214] Hum. Мол. Genet. 1994; 3: 421–428. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Hayes JD, Strange RC. Полиморфизмы глутатион-S-трансферазы и их биологические последствия. Фармакология. 2000. 61: 154–166. [PubMed] [Google Scholar]
- Ishikawa K, Funayama T, Ohde H, Inagaki Y, Mashima Y. Генетические варианты TP53 и EPHX1 при наследственной оптической невропатии Лебера и их связь с возрастом в начале болезни.Jpn. J. Ophthalmol. 2005. 49: 121–126. [PubMed] [Google Scholar]
- Ланкастер Дж. М., Браунли Х.А., Белл Д.А., Футреал ПА, Маркс Дж. Р., Берчак А., Вайзман Р. У., Тейлор Дж. А. Полиморфизм микросомальной эпоксидгидролазы как фактор риска рака яичников. Мол. Канцерогенный. 1996. 17: 160–162. [PubMed] [Google Scholar]
- Леконт Т., Ланди Б., Бон П., Лоран-Пуч П., Лорио М.-А. Полиморфизм глутатион-S-трансферазы P1 (Ile105Val) предсказывает кумулятивную невропатию у пациентов, получающих химиотерапию на основе оксалиплатина.Clin. Cancer Res. 2006; 12: 3050–3056. [PubMed] [Google Scholar]
- Линд Дж., Тондель М., Эстерберг А., Вретем М. Криптогенная полинейропатия: клинические и нейрофизиологические данные. Дж. Перифер. Nerv. Syst. 2005; 10: 31–37. [PubMed] [Google Scholar]
- Маннервик Б. Изоферменты глутатионтрансферазы. Adv. Энзимол. Relat. Районы Мол. Биол. 1985; 57: 357–417. [PubMed] [Google Scholar]
- Martyn CN, Hughes RA. Эпидемиология периферической невропатии. J. Neurol. Нейрохирургия.Психиатрия. 1997. 62: 310–318. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Оеш Ф. Эпоксидные гидразы млекопитающих: индуцируемые ферменты, катализирующие инактивацию канцерогенных и цитотоксических метаболитов, полученных из ароматических и олефиновых соединений. Xenobiotica. 1973; 3: 305–340. [PubMed] [Google Scholar]
- Omiecinski CJ, Aicher L, Holubkov R, Checkoway H. Периферические лимфоциты человека как индикаторы активности микросомальной эпоксидгидролазы в печени и легких. Фармакогенетика. 1993; 3: 150–158.[PubMed] [Google Scholar]
- Реббек TR. Молекулярная эпидемиология генотипов GSTM1 и GSTT1 глутатион-S-трансферазы человека в отношении предрасположенности к раку. Cancer Epidemiol. Биомаркеры Пред. 1997; 6: 733–743. [PubMed] [Google Scholar]
- Шатунов А., Мок К., Ньюхаус С., Уил М. Е., Смит Б., Вэнс С., Джонсон Л., Велдинк Дж. Х., ван Эс М. А., ван ден Берг Л. Х. и др. Хромосома 9p21 при спорадическом боковом амиотрофическом склерозе в Великобритании и семи других странах: исследование ассоциации на уровне всего генома.Lancet Neurol. 2010; 9: 986–994. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Smith CAD, Harrison DJ. Связь между полиморфизмом гена микросомальной эпоксидгидролазы и восприимчивостью к эмфиземе. Ланцет. 1997; 350: 630–633. [PubMed] [Google Scholar]
- Седерквист П., Ахмади А., Акербак А., Аксельсон О., Флодин Ю. Нулевой генотип M1 глутатион-S-трансферазы как модификатор риска хронической токсической энцефалопатии, вызванной растворителем. Скандинавский журнал работы, окружающей среды и здоровья.1996. 22: 360–363. [PubMed] [Google Scholar]
- Spencer PS, Schaumburg HH. Экспериментальная и клиническая нейротоксикология. Оксфорд: Oxford Univ. Нажмите; 2000. [Google Scholar]
- Spencer PS, Sabri MI, Schaumburg HH, Moore CL. Является ли дефект энергетического обмена нервного волокна причиной дегенерации аксонов при полинейропатиях? Аня. Neurol. 1979; 5: 501–507. [PubMed] [Google Scholar]
- Tesfaye S, Chaturvedi N, Eaton SE, Ward JD, Manes C, Ionescu-Tirgoviste C, Witte DR, Fuller JH.Факторы сосудистого риска и диабетическая невропатия. N. Engl. J. Med. 2005; 352: 341–350. [PubMed] [Google Scholar]
- Teunissen LL, Franssen H, Wokke JH, van der Graaf Y, Linssen WH, Banga JD, Laman DM, Notermans NC. Являются ли сердечно-сосудистые заболевания фактором риска развития аксональной полинейропатии? J. Neurol. Нейрохирургия. Психиатрия. 2002; 72: 590–595. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Тимбрелл Дж. Факторы, влияющие на токсические реакции: метаболизм. В: Фрэнсис Т., редактор. Принципы биохимической токсикологии.Лондон: Taylor & Francis Ltd .; 2000. С. 65–110. [Google Scholar]
- Тондель М., Линд Дж., Йонссон П., Вретем М., Перссон Б. Профессиональные детерминанты криптогенной полинейропатии. Нейроэпидемиология. 2006; 26: 187–194. [PubMed] [Google Scholar]
- Сюй С., Ван И, Роу Б., Пирсон В. Р.. Характеристика генного кластера глутатион-S-трансферазы человеческого класса и делеции GSTM1. J. Biol. Chem. 1998. 273: 3517–3527. [PubMed] [Google Scholar]
- Чжан Ю., Лю Цюй, Дуань Х, Ченг Дж, Цзян С., Хуанг Х, Ленг С., Хэ Ф, Чжэн Ю.Связь между полиморфизмом метаболических генов и восприимчивостью к повреждению периферических нервов у рабочих, подвергшихся воздействию н-гексана: предварительное исследование. Биомаркеры. 2006; 11: 61–69. [PubMed] [Google Scholar]
Требования — цифровые электронные системы
Требования к общеобразовательному | Часы | |
КУ Ядро 2.1 | (обычно ENGL 101 и ENGL 102) | 6 |
Химия | CHEM 130, Основы химии I, или CHEM 150 Химия для инженеров, или CHEM 190 Основы химии I, с отличием вместе с лабораторией CHEM 191, основы химии I, с отличием. | 5 |
KU Основные требования * | Курсы, утвержденные для достижения целей 2.2, 3S, 3N, 4.1 и 4.2. Обратите внимание, что PHSX 594 соответствует цели 4.2. | 15 |
Физика ядра | Часы | |
PHSX 150 | Семинар по физике, астрономии и инженерной физике | 0.5 |
PHSX 211 и 216 или PHSX 213 | Лаборатория общей физики I и общей физики I (213 — это диплом с отличием / специальностью) | 5 |
PHSX 212 и 236 или PHSX 214 | Лаборатория общей физики II и общей физики II (214 — это эквивалент диплома с отличием / специальностью) | 4 |
PHSX 313 | Общая физика III | 3 |
PHSX 316 | Лаборатория промежуточной физики (с или после PHSX 313) | 1 |
EPHX 516 | Физические измерения | 4 |
EPHX 521 | Механика I | 3 |
EPHX 531 | Электричество и магнетизм | 3 |
EPHX 601 | Проектирование физических и электронных систем | 4 |
Математика | Часы | |
MATH 125 | Исчисление I (MATH 145 — эквивалент с отличием) | 4 |
MATH 126 | Calculus II (MATH 146 эквивалентно с отличием) | 4 |
MATH 127 | Calculus III (MATH 147 эквивалентно с отличием) | 4 |
MATH 290 | Элементарная линейная алгебра (MATH 291 эквивалентно с отличием) | 2 |
MATH 220 или 320 | Прикладные или элементарные дифференциальные уравнения.(MATH 221 является эквивалентом с отличием) | 3 |
Особые требования к вариантам конструкции в цифровых электронных системах | Часы | |
Физика | EPHX 511, Введение в квантовую механику | 3 |
Математика | MATH 526 Прикладная математическая статистика или MATH 628 1 Математическая теория статистики | 3 |
Электротехника и Компьютерные науки | EECS 140, Введение в дизайн цифровой логики (EECS 141 — эквивалент с отличием) | 4 |
EECS 168, Программирование I (EECS 169 — эквивалент с отличием) | 4 | |
EECS 211, Цепи I | 3 | |
EECS 212, Цепи II | 4 | |
EECS 268, Программирование II | 4 | |
EECS 312, Электронные схемы I. | 3 | |
EECS 360, Анализ сигналов и систем | 4 | |
EECS 388, Компьютерные системы и язык ассемблера | 4 | |
EECS 443, Проектирование цифровых систем | 4 | |
EECS 448, Разработка программного обеспечения I | 4 | |
EECS 470, Электронные устройства и свойства материалов | 3 | |
EECS 541, Лаборатория проектирования компьютерных систем I | 3 | |
EECS 542, Лаборатория проектирования компьютерных систем II | 3 | |
EECS 645 Архитектура компьютера | 3 | |
EECS, факультатив | 3 | |
1 MATH 628 предлагается только весной и требует MATH 627 в качестве предварительного условия.
* Цель 1.1, цель 1.2, цель 3N, цель 5 и цель 6 удовлетворяют основным требованиям.
Требования к общеобразовательному | Часы | |
КУ Ядро 2.1 | (обычно ENGL 101 и ENGL 102) | 6 |
Химия | CHEM 130, Основы химии I или CHEM 150, Химия для инженеров | 5 |
KU Основные требования | курсов, утвержденных для достижения Целей 2.2, 3S, 3N, 4.1 и 4.2 | 15 |
Физика ядра | Часы | |
PHSX 150 | Семинар по физике, астрономии и инженерной физике | 0,5 |
PHSX 211/216 или 213 | Лаборатория общей физики I и общей физики I (213 — это диплом с отличием / специальностью) | 5 |
PHSX 212/236 или 214 | Лаборатория общей физики II и общей физики II (214 — это эквивалент диплома с отличием / специальностью) | 4 |
PHSX 313 | Общая физика III | 3 |
PHSX 316 | Лаборатория промежуточной физики (с или после PHSX 313) | 1 |
EPHX 516 | Физические измерения | 4 |
EPHX 521 | Механика I | 3 |
EPHX 531 | Электричество и магнетизм | 3 |
PHSX 601 | Проектирование физических и электронных систем | 4 |
Математика | Часы | |
MATH 125 | Исчисление I | 4 |
MATH 126 | Исчисление II | 4 |
MATH 127 | Исчисление III | 4 |
MATH 290 | Элементарная линейная алгебра | 2 |
MATH 220 или 320 | Прикладные или элементарные дифференциальные уравнения | 3 |
Особые требования к вариантам конструкции в цифровых электронных системах | Часы | |
Физика | EPHX 511, Введение в квантовую механику | 3 |
Избран.Engr & | EECS 140, Введение в цифровой логический дизайн (или EECS 141, с отличием) | 4 |
Computer Sci. | EECS 168, Программирование I | 4 |
EECS 211, Цепи I | 3 | |
EECS 212, Цепи II | 4 | |
EECS 268, Программирование II | 4 | |
EECS 312, Электронные схемы I. | 3 | |
EECS 360, Анализ сигналов и систем | 4 | |
EECS 388, Компьютерные системы и язык ассемблера | 4 | |
EECS 443, Проектирование цифровых систем | 4 | |
EECS 448, Разработка программного обеспечения I | 4 | |
EECS 461, Вероятность и статистика | 3 | |
EECS 470, Электронные устройства и свойства материалов | 3 | |
EECS 541, Лаборатория проектирования компьютерных систем I | 3 | |
EECS 542, Лаборатория проектирования компьютерных систем II | 3 | |
EECS 645 Архитектура компьютера | 3 | |
EECS, факультатив | 3 | |
KU Основная учебная программа и требования по химии (26 кредитов) | Часы | |
Английский | Результат 1 цели 2 основной учебной программы KU | 6 |
Химия | CHEM 130, Основы химии, или CHEM 150, Химия для инженеров, или CHEM 170, Химия для химических наук I | 5 |
KU Основные требования | Выполнение требований основной учебной программы Цели 2, Результат 2, Цели 3 (искусство и гуманитарные науки, социальные науки) и Цели 4, результаты 1 и 2. | 15 |
Основные курсы физики (27,5 кредита) | Часы | |
PHSX 150 | Семинар по физике, астрономии и инженерной физике | 0,5 |
PHSX 211 или 213 | Общая физика I (213 — эквивалент диплома с отличием, 5 баллов, включая лабораторную работу) | 4 |
PHSX 212 или 214 | Общая физика II (214 — эквивалент диплома с отличием, 4 кредита, включая лабораторную работу) | 3 |
PHSX 216 | Лаборатория общей физики I | 1 |
PHSX 236 | Лаборатория общей физики II | 1 |
PHSX 313 | Общая физика III | 3 |
PHSX 316 | Лаборатория промежуточной физики (с или после PHSX 313) | 1 |
EPHX 516 | Физические измерения | 4 |
EPHX 521 | Механика I | 3 |
EPHX 531 | Электричество и магнетизм | 3 |
PHSX 601 | Проектирование физических и электронных систем | 4 |
Курсы математики (18 кредитов) | Часы | |
MATH 121 | Исчисление I | 5 |
MATH 122 | Исчисление II | 5 |
MATH 223 | Векторное исчисление | 3 |
MATH 290 | Элементарная линейная алгебра | 2 |
MATH 220 | Прикладные или элементарные дифференциальные уравнения | 3 |
Требования к курсу для варианта дизайна в цифровых электронных системах (59 кредитов) | Часы | |
Физика | EPHX 511, Введение в квантовую механику | 3 |
Избран.Engr & | EECS 140, Введение в цифровой логический дизайн (или EECS 141, с отличием) | 4 |
Computer Sci. | EECS 168, Программирование I | 4 |
EECS 211, Цепи I | 3 | |
EECS 212, Цепи II | 4 | |
EECS 268, Программирование II | 4 | |
EECS 312, Электронные схемы I. | 3 | |
EECS 360, Анализ сигналов и систем | 4 | |
EECS 388, Компьютерные системы и язык ассемблера | 4 | |
EECS 443, Проектирование цифровых систем | 4 | |
EECS 448, Разработка программного обеспечения I | 4 | |
EECS 461, Вероятность и статистика | 3 | |
EECS 470, Электронные устройства и свойства материалов | 3 | |
EECS 541, Лаборатория проектирования компьютерных систем I | 3 | |
EECS 542, Лаборатория проектирования компьютерных систем II | 3 | |
EECS 645 Computer Architecture или 643, Advanced Computer Architecture | 3 | |
EECS Elective | 3 | |
Систематический подход к анализу взаимодействия генов: полиморфизмы в микросомальной эпоксидгидролазе EPHX и глутатион-S-трансферазе GSTM1, GSTT1 и GSTP1 и риск рака молочной железы
Abstract
Цель: Мы провели исследование случай-контроль в выборке населения европеоидной расы Австралии, состоящей из 1246 случаев и 664 контролей, для оценки роли полиморфизма гена детоксикации EPHX T> C Tyr 113 His, GSTT1 делеция, Делеция GSTM1 и GSTP1 A> G Ile 105 Val на риск рака груди.
Методы: мы систематически обращались к основным эффектам и возможным взаимодействиям генов с использованием безусловной логистической регрессии для оценки отношения шансов (OR) с поправкой на потенциальные искажающие факторы и с использованием стандартных подходов к построению моделей, основанных на теории правдоподобия.
Результаты. Было отмечено снижение риска, связанного с генотипом EPHX CC [OR, 0,60; 95% доверительный интервал (95% ДИ) 0,43-0,84; P = 0,003], незначительно значимое свидетельство повышенного риска с нулевым генотипом GSTM1 (OR, 1.21; 95% ДИ, 1,00–1,47; P = 0,05), но нет ассоциации с GSTT1 нулевым генотипом (OR, 1,12; 95% ДИ, 0,86-1,45; P = 0,4) или GSTP1 (OR, 0,95; 95% ДИ, 0,82- 1.10; P = 0.5) генотип. Полная модель со всеми взаимодействиями дала значительно лучшее соответствие, чем модель только с основными эффектами ( P <0,001), обеспечивая доказательства взаимодействия ген-ген. Самая экономная модель включала основные эффекты для EPHX, GSTT1 и GSTM1 ; двустороннее взаимодействие между EPHX и GSTM1 ; и трехстороннее взаимодействие между EPHX, GSTM1 и GSTT1 .Прогнозируемые риски были наибольшими для женщин с делециями GSTT1 и GSTM1 , либо с генотипом EPHX TC (OR, 2,02; 95% ДИ, 1,19-3,45; P = 0,009), либо с EPHX CC . генотип (OR, 3,54; 95% ДИ, 1,29-9,72; P = 0,14).
Заключение: Полиморфизмы генов детоксикации могут взаимодействовать друг с другом, в результате чего небольшие группы людей подвергаются умеренно повышенному риску. Мы предостерегаем от чрезмерной интерпретации и предполагаем, что необходимо объединение таких же крупных исследований, чтобы прояснить возможную роль таких сложных взаимодействий генов в риске рака груди.(Биомаркеры эпидемиологии рака, пред. 2007; 16 (4): 769–74)
- рак груди
- GST гены
- EPHX
- полиморфизмы
Введение
Генетические факторы риска рака груди включают доминантно наследуемые вредные мутации в генах BRCA1 и BRCA2 . Однако носители таких мутаций редки на уровне популяции и составляют небольшую часть семейной агрегации, наблюдаемой при раке груди (1).Таким образом, возможно, что более часто встречающиеся генетические факторы «низкого риска» объясняют значительную часть остаточной семейной агрегации (2). Утверждалось, что эти эффекты могут быть очевидны только при рассмотрении совместного действия нескольких генетических факторов и факторов окружающей среды, которые действуют в рамках биологического пути (3), хотя поиск этих взаимодействий требует некоторой осторожности, чтобы уменьшить частоту ложноположительных выводов. (4).
Гены-кандидаты предрасположенности к раку груди включают те, которые участвуют в метаболизме канцерогенов, содержащих аллельные варианты, влияющие на экспрессию генов или функцию белков.К ним относятся ген микросомальной эпоксидгидролазы фазы I EPHX и гены глутатиона S -трансферазы фазы II GSTT1, GSTM1 и GSTP1. EPHX кодирует белок эпоксидгидролазы mEH, который катализирует гидролиз арена и алифатических эпоксидов до транс- -дигидродиолов (5) и обычно приводит к детоксикации и подготовке к реакциям конъюгации фазы II. Гены GST катализируют опосредованное глутатионом восстановление экзогенных и эндогенных электрофилов с широкой и перекрывающейся субстратной специфичностью (6, 7), обычно производя легко выводимые водорастворимые соединения.Таким образом, аллельные варианты, связанные с измененными (более быстрыми или медленными) темпами детоксикации потенциальных канцерогенов, могут повышать восприимчивость к раку, возможно, в большей степени при наличии стрессов окружающей среды, таких как курение и воздействие УФ-излучения (8).
Существует ряд функциональных вариантов-кандидатов для генов EPHX и GST . Было показано, что экзон 3 EPHX T до C Tyr 113 His-вариант с аминокислотной заменой имеет функциональное значение in vitro (9), причем вариант His 113 демонстрирует 60% активности относительно дикий тип Tyr 113 .Вариант с делецией или нулевые аллели существуют для генов GSTT1 и GSTM1 (10-12) и биохимически проявляются как неспособность экспрессировать белок. Эти аллели являются общими, и нулевые генотипы GSTT1 и GSTM1 встречаются у от ~ 10% до 20% и 50% населения европеоидной расы, соответственно (13). GSTP1 экзон 5 A — G Ile 105 Val полиморфизм является одним из нескольких вариантов, описанных для этого локуса (14, 15), и было показано, что Val-содержащая изоформа изменила удельную активность и уменьшила тепло стабильность (16-18).
Эти и другие полиморфизмы в вышеупомянутых генах были исследованы как факторы риска многих видов рака, независимо или в комбинации. Результаты отдельных исследований случай-контроль, оценивающих влияние полиморфизма GST на риск рака груди, противоречивы. Два метаанализа (см. Обсуждение) обнаружили значительную связь рака груди с генотипом делеции GSTM1 у женщин в постменопаузе (19, 20), с более сильными доказательствами и более высокими отношениями шансов (OR) из исследований в группах населения с низкой частотой. нулевого генотипа GSTM1 (20).В одном исследовании сообщалось о небольшом, незначительно значимом увеличении риска, связанного с генотипом с делецией GSTT1 (19). Однако недавний объединенный анализ (см. «Обсуждение») Vogl et al. (21) не обнаружили значимой связи с общим риском рака груди для GSTT1 или GSTM1 как для женщин в пременопаузе, так и в постменопаузе. Мета-анализ GSTP1 экзона 5 A> G Ile 105 Val полиморфизма не выявил доказательств того, что этот вариант был связан с риском либо для гетерозиготного генотипа (21), либо для гомозиготного генотипа (19, 21).Кроме того, в объединенном анализе (21) не было доказательств того, что генотипы GSTT1, GSTM1 или GSTP1 действовали в комбинации для увеличения риска рака груди или взаимодействовали с курением или репродуктивным анамнезом, чтобы изменить риск рака груди.
В единственном опубликованном исследовании воздействия EPHX на рак груди (22) сообщается о незначительном повышении риска в 1,5 раза, связанном с генотипом EPHX экзон 3 CC по сравнению с генотипом TT дикого типа и пограничный значительный 2.В 2 раза повышен риск для EPHX экзона 3, CC генотипа в сочетании с GSTM1 нулевым генотипом. Однако это больничное исследование с участием в основном женщин в постменопаузе было относительно небольшим и включало 238 случаев и 313 контрольных.
Ввиду неопределенности в литературе мы провели исследование случай-контроль в Австралии, чтобы оценить роль генетических полиморфизмов GSTT1, GSTM1, GSTP1 и EPHX как факторов риска рака груди.Мы специально провели систематический анализ возможных взаимодействий между этими генетическими полиморфизмами.
Материалы и методы
Субъектов
Австралийское семейное исследование рака молочной железы, популяционное исследование рака груди у женщин в возрасте до 60 лет с использованием метода «случай-контроль», было проведено в Мельбурне и Сиднее с упором на раннее начало заболевания (23). Детали методов сбора данных были описаны ранее (23, 24).Вкратце, это случаи, когда у женщин был диагностирован первый первичный рак груди, выявленный в реестрах рака Виктории и Нового Южного Уэльса, а контрольными случаями были женщины, ранее не имевшие рака груди, выбранные из списков избирателей (регистрация взрослых для голосования в Австралии обязательна) стратифицированная случайная выборка, подобранная по возрасту. Анкеты использовались для измерения факторов риска; семейный анамнез рака систематически собирался для каждого случая и контролировался из нескольких источников в семье; и образцы крови были собраны во время интервью у субъектов, согласившихся участвовать в генетических исследованиях.Генотипирование проводилось на основе наличия ДНК. Одобрение на исследование было получено от этических комитетов Мельбурнского университета, Онкологических советов Виктории и Нового Южного Уэльса и Института медицинских исследований Квинсленда. Все субъекты предоставили письменное информированное согласие.
Лица, включенные в генетический анализ (86% участвующих субъектов), отличались от остальных участвующих субъектов в отношении определенных факторов, которые ранее были связаны с раком груди (25), а именно семейного анамнеза (определяемого как наличие по крайней мере одного первого или родственник второй степени родства с раком груди), использование оральных контрацептивов и равенство.
Этническая принадлежность была указана самостоятельно. Субъекты, имеющие какое-либо происхождение австралийских аборигенов, жителей островов Торресова пролива или маори; или страны рождения в южной части Тихого океана, Индийского океана, Карибских островов или Азии были классифицированы как некавказские. Предварительный анализ показал, что распределение генотипов для GSTT1, GSTP1 и EPHX различается между людьми, сообщающими об этнической принадлежности европеоидной и некавказской национальности, что согласуется с данными других исследований (см. Раздел «Обсуждение»). Таким образом, дальнейший анализ был ограничен участниками, которые идентифицировали себя как европеоид.Кроме того, молекулярные анализы на сегодняшний день (24) 5 выявили 59 случаев европеоидной расы, несущих вредную мутацию зародышевой линии в BRCA1 или BRCA2 , и эти участники также были исключены из анализа. Окончательный анализ включал 1246 случаев и 664 контроля.
Измерение генотипа
Генотипирование было, как описано ранее (26, 27). Вкратце, методология ПЦР-агарозы была использована для обнаружения генотипов с гомозиготной делецией GSTT1 и GSTM1 , а методология системы определения последовательности Taqman с призмой ABI 7700 была использована для генотипирования EPHX T — C His полиморфизм (rs1051740) и GSTP1 A от до G Ile 105 Val полиморфизм (rs1695).Кроме того, поскольку предварительный анализ показал отклонение от равновесия Харди-Вайнберга, все индивидуумы с EPHX CC и подгруппа индивидуумов с генотипом TC были регенотипированы с использованием стандартной денатурирующей высокоэффективной жидкостной хроматографии и методологии секвенирования. Последовательности праймеров были следующими: прямая, GCTTCCACTATGGCTTC и обратная, TTGGGTTCTGAATCTCTCCAA, и ПЦР проводили с использованием программы приземления с конечной температурой отжига 55 ° C и температурой плавления при денатурирующей высокоэффективной жидкостной хроматографии 59 ° C.Эту методологию использовали для подтверждения генотипа в позиции Tyr 113 His, а также для установления того, что никакие другие вариации, такие как описанный вариант Trp 97 Stop, не могут отрицательно сказаться на результатах анализа. Подтверждение последовательности всех аберрантных денатурирующих профилей высокоэффективной жидкостной хроматографии, обнаруженных во время этого повторного скрининга, не выявило никаких доказательств дополнительных вариаций под праймером Taqman и сайтами связывания зонда, а также идентифицировало ошибочную классификацию генотипа в 0,5% случаев и 0.2% контрольных, с повторным вызовом генотипа CC для генотипирования Taqman.
Статистический анализ
Равновесие Харди-Вайнберга проверялось с помощью теста Пирсона χ 2 . Связь статуса случай-контроль с генотипом и факторами окружающей среды моделировалась с помощью безусловной логистической регрессии. EPHX и GSTP1 изначально моделировались как кодоминантные генотипы. Впоследствии EPHX и GSTP1 были включены как линейные термины, закодированные 0, 1 или 2, с гомозиготным диким типом ( TT для EPHX, AA для GSTP1 ), закодированным как 0.Тест отношения правдоподобия использовался для оценки любого улучшения соответствия между моделью кодоминантного генотипа и линейной моделью для EPHX и GSTP1. GSTM1 и GSTT1 каждый был закодирован таким образом, чтобы гомозиготная делеция контрастировала с объединенными гетерозиготными и дикими типами, что отражало чувствительность используемых методов генотипирования на основе геля.
В дополнение к генотипу, был также включен ряд факторов окружающей среды, которые считаются потенциально искажающими: эталонные возрастные категории (≤30, 31-40, 41-50,> 50 лет), статус менопаузы, квартиль возраста наступления менархе, количество живорождений (0, 1, 2, 3, ≥4), семейный анамнез (любой родственник первой или второй степени родства, у которого был рак груди), предшествующее доброкачественное заболевание груди, индекс массы тела [вес (кг) / рост (m) 2 ] квартиль, курение (никогда / никогда), регулярное употребление алкоголя, использование оральных контрацептивов (никогда / никогда), заместительная гормональная терапия когда-либо, образование (три категории), страна рождения и текущее состояние Австралии. резиденция.
Систематическая процедура построения модели использовалась для включения или исключения генетических полиморфизмов и их взаимодействий с использованием теста отношения правдоподобия для расчета статистической значимости вложенных моделей (28). Использовались как прямой отбор, так и обратное исключение. Помимо основных эффектов EPHX, GSTT1, GSTM1 и GSTP1 , мы рассмотрели все возможные двух-, трех- и четырехсторонние взаимодействия ген-ген. Каждая генетическая модель рассматривалась без включения каких-либо факторов окружающей среды, включая все перечисленные выше, и включая только те, которые оставались значимо связанными согласно тесту Вальда.Из-за роли этих генов-кандидатов в детоксикации мы также рассмотрели взаимодействие каждого аллеля с курением и регулярным употреблением алкоголя.
Результаты были представлены в виде решетки, показывающей для каждой модели статистику G , определяемую как удвоенную разницу в логарифмических вероятностях между подобранной моделью и нулевой моделью (следовательно, G для нулевой модели = 0 ). Первый уровень показывает нулевую модель, следующий уровень показывает модели с 1 степенью свободы ( df ), следующий с 2 df и так далее.Каждый уровень рассматривает все возможные комбинации переменных. Поскольку включение эффекта для GSTP1 не имело существенного значения ни для одной из подобранных моделей (см. Результаты ниже), мы исключили его из решетки, чтобы упростить представление (см. Рис. 1 ). Различия в статистике G между вложенными моделями сравниваются с распределением χ 2 с df = разница в df вложенных моделей.
Рисунок 1.Модель строительной решетки. A, EPHX ; B, GSTM1 ; C, GSTT1 ; подчеркнутых, условий взаимодействия. Тест отношения правдоподобия: количество переменных ( df ) в моделях увеличивается с каждой строкой. Различия в G = 2 ln (вероятность подобранной модели / вероятность нулевой модели) между вложенными моделями (, соединенными линиями ) можно сравнить с распределением χ 2 с df = разница модели df . Обратите внимание, что генотип GSTP1 не был связан с раком груди, и добавление терминов, относящихся к этому генотипу, не привело к более чем минимальному изменению любых других оценок.Поэтому для простоты GSTP1 не показан на диаграмме решетки.
Всякий раз, когда были обнаружены доказательства взаимодействия генов с риском рака, мы тестировали независимый ассортимент отдельно в случаях и в контролях с использованием лог-линейной модели (29). Тот же подход использовался для проверки связи между генотипом и курением, а также между генотипом и регулярным употреблением алкоголя. Наконец, анализы были повторены без учета факторов окружающей среды. После оценки окончательных моделей корреляционная матрица оценок была проверена на наличие коллинеарности.Уровень значимости 5% был выбран априори, и все тесты были двусторонними. Для анализа использовалась программа Stata (версия 9.0, StataCorp).
Результаты
Распределение генотипов для GSTT1, GSTP1 и EPHX различается между людьми, сообщающими об этнической принадлежности европеоидной и некавказской национальности ( P <0,001; P = 0,013; P = 0,002, соответственно). Таким образом, дальнейшие анализы были ограничены женщинами, которые идентифицировали себя как европеоид.Это включало 663 контроля и 1238 случаев, генотипированных по EPHX ; 649 контролей и 1232 случая генотипирования по GSTP1 ; 664 контроля и 1246 случаев генотипирования по GSTT1 и GSTM1 , соответственно (Таблица 1 ).
Таблица 1.Необработанные и скорректированные одномерные OR для ассоциации между раком груди и генотипами EPHX, GSTP1, GSTM1 и GSTT1
Признаки отклонения от равновесия Харди-Вайнберга в контроле (меньше гетерозигот, чем ожидалось) наблюдались для EPHX и сохранялись после подтверждения генотипов с использованием альтернативной методологии генотипирования ( P = 0.01). Не было доказательств отклонения от равновесия Харди-Вайнберга в контроле для GSTP1 ( P = 0,6).
Ни в одном случае не было обнаружено существенных различий между нескорректированными и скорректированными оценками. Скорректированные оценки представлены в тексте. Как показано в таблице 1, значительное снижение риска рака груди было связано с генотипом EPHX CC по сравнению с генотипом TT [OR, 0,60; 95% доверительный интервал (95% ДИ), 0.43-0,84; P = 0,003; OR тренда 0,83; 95% ДИ 0,72-0,96; P = 0,01]. Связи риска рака груди с генотипом GSTP1 не было ( GG по сравнению с AA OR, 0,90; 95% ДИ, 0,66–1,25; OR тенденции, 0,95; 95% ДИ, 0,82–1,10). Кодоминантная модель для EPHX и GSTP1 не обеспечила значительно лучшего соответствия, чем линейная модель ( P = 0,1 и P = 0,9, соответственно). Имеются незначительно значимые доказательства повышенного риска рака груди с нулевым генотипом GSTM1 (OR, 1.21; 95% ДИ, 1,00–1,47; P = 0,05), но не с нулевым генотипом GSTT1 (OR, 1,12; 95% ДИ, 0,86-1,45; P = 0,4).
Было обнаружено незначительное взаимодействие между употреблением алкоголя (никогда / никогда) и GSTP1 (OR взаимодействия, 1,36; 95% ДИ, 1,01-1,83; P = 0,04) и предполагаемая связь между курением (никогда / никогда) и GSTP1 (OR взаимодействия, 1,30; 95% ДИ, 0,97–1,73; P = 0,09). Поскольку слабое взаимодействие могло быть результатом ассоциации между поведением и генотипом на уровне популяции, эти ассоциации были протестированы только для контрольной группы и оказались несущественными для курения (OR, 0.88; 95% ДИ, 0,65–1,21; P = 0,4) и питьевой (OR 0,83; 95% ДИ 0,61–1,15; P = 0,3). Взаимодействие между генотипом и статусом менопаузы и другими репродуктивными факторами, такими как половая принадлежность и возраст наступления менархе, также не было значимым (данные не показаны).
Результаты как прямого отбора, так и обратного исключения исключили все основные эффекты и взаимодействия ген-ген с GSTP1 ( P от 0,3 до 0,95). Поэтому для простоты необработанные данные для случаев и элементов управления в Таблице 2 , и результаты процедуры построения модели, показанной на рис.1, был показан без GSTP1 .
Таблица 2.Количество случаев и контролей по GSTM1, GSTT1 и Статус генотипа EPHX
Модель только с основными эффектами дала значительно лучшее соответствие, чем нулевая модель (χ 2 = 9,94 на 3 df ; P <0,02), даже после добавления незначительного эффекта для GSTP1 ( P <0,05; данные не показаны). Полная модель также дала значительно улучшенное соответствие по сравнению с нулевой моделью (χ 2 = 31.15 по 7 дф ; P <0,0001), даже после добавления восьми незначительных эффектов для всех взаимодействий с GSTP1 ( P <0,01; данные не показаны). Полная модель также дала значительно лучшее соответствие, чем модель только с основными эффектами (χ 2 = 21,21 на 4 df ; P <0,001), даже с учетом GSTP1 ( P <0,02 ). Таким образом, были доказательства наличия взаимодействия ген-ген.
Наиболее экономная подгонка была обеспечена выделенной моделью на рис. 1 с 5 df , с G = 30,26. Подгонка была значительно лучше, чем у лучших моделей с 4 df ( P = 0,02), 2 df ( P = 0,003) или 1 df ( P = 0,0001). Лучшая модель 3 df не была вложена в самую экономную модель и не может быть формально сравнена, но разница AIC в 11,2 предполагает, что окончательная модель дала лучшее соответствие.Более сложные модели не дали лучшего соответствия: лучший 6 df дал G 6 = 30,55 ( P = 0,6), тогда как модель 7 df со всеми возможными взаимодействиями дала G 7 = 31,15 ( P = 0,6).
Как описано в таблице 3 наиболее экономная модель, которая возникла в результате этой процедуры, состояла из основных эффектов для EPHX (OR, 0,92; P = 0,5), GSTT1 (OR, 0.85; P = 0,3) и GSTM1 (OR, 1,36; P = 0,02), двустороннее взаимодействие между EPHX и GSTM1 (взаимодействие OR, 0,72; P = 0,03), и трехстороннее взаимодействие между EPHX, GSTM1 и GSTT1 (OR взаимодействия, 2,63 на аллель C EPHX , P = 0,001). Это подразумевает OR 2,02 (95% ДИ 1,19–3,45; P = 0,009) для тех, у кого нет действующей копии GSTM1 или GSTT1 и генотипа TC для EPHX .В сочетании с генотипом EPHX CC это OR становится 3,54 (95% ДИ, 1,29–9,72; P = 0,01). Когда GSTP1 был добавлен обратно в окончательную модель в качестве основного эффекта, не было ни доказательств ассоциации в качестве основного эффекта (OR, 0,95; P = 0,5), ни модификации эффектов других генов (результаты не показаны. ).
Таблица 3.Оценки основных эффектов и эффектов взаимодействия для EPHX, GSTM1 и GSTT1 из наиболее экономичной модели
Когда наиболее экономная модель была приспособлена к случаям и контролю отдельно с помощью регрессии Пуассона, двустороннее взаимодействие было значимым только для случаев ( P = 0.035). Не было доказательств независимого ассортимента EPHX и GSTM1 для контроля ( P = 0,2). Однако казалось, что существует трехсторонняя связь между EPHX, GSTT1 и GSTM1 для элементов управления. Хотя эта зависимость также наблюдалась в случаях, ассоциация была больше для контроля (оценочный коэффициент = -0,6; P = 0,002), чем для случаев (оценочный коэффициент = 0,2; P = 0,05).
Обсуждение
Гены детоксикации долгое время считались вероятными генами-кандидатами на предрасположенность к раку груди и другим видам рака. Наше исследование случай-контроль выборки рака молочной железы с относительно ранним началом выявило доказательства защитного эффекта, связанного с генотипом EPHX CC , наводящие на размышления доказательства повышенного риска, связанного с генотипом с делецией GSTM1 , и отсутствие связи с генотипом . Генотип с делецией GSTT1 или вариантный генотип GSTP1 в целом.Эти ассоциации не были изменены менопаузальным статусом или другими факторами репродуктивного риска, и не было убедительных доказательств взаимодействия с курением сигарет или регулярным употреблением алкоголя.
Поскольку эти гены действуют по общему пути, мы систематически априори тестировали взаимодействия ген-ген, используя иерархическую статистическую модель. Мы обнаружили доказательства того, что добавление взаимодействий ген-ген дало лучшее соответствие, чем только основные эффекты, поскольку модели, которые включали взаимодействия, давали значительно лучшее соответствие, чем модель, включающая только основные эффекты.Из всех моделей взаимодействия самая экономная предсказывала, что делеции GSTM1 и GSTT1 были связаны с двукратным повышением риска рака груди при совместном наследовании с генотипом EPHX TC и в 3,5 раза с генотипом EPHX TC. CC генотип.
Участие GST генов в этиологии рака молочной железы было широко изучено в большом количестве небольших исследований с противоречивыми результатами, и совместный анализ опубликованных наборов данных не дал твердых выводов относительно их роли в риске рака молочной железы.Мета-анализ не менее 2000 случаев и 2000 контрольных исследований из 10-15 исследований преимущественно женщин европеоидной расы, проведенный Egan et al. (19) обнаружили погранично значимую связь с GSTM1 только у женщин в постменопаузе (OR, 1,14), пограничное значимое увеличение риска делеции GSTT1 (OR, 1,11) и отсутствие связи с генотипом GSTP1 GG (OR , 1.04). Мета-анализ данных GSTM1 из более чем 2000 случаев и 2000 контрольных данных из 10 исследований Sull et al.(20) также не обнаружили общей ассоциации для генотипа с делецией, но значительно увеличили риск только для женщин в постменопаузе (объединенное ОШ, 1,19), особенно для популяций с низкой частотой нулевого аллеля GSTM1 . Меньший объединенный анализ Vogl et al. (21) из семи исследований случай-контроль (558–1899 контрольных, 921–2033 случаев) не обнаружили связи для GSTM1 (OR, 0,98), GSTT1 (OR, 1.11) или GSTP1 (OR для CC генотип 0,93) и отсутствие взаимодействия с курением, гормональными факторами риска или комбинациями этих генов.Наконец, метаанализ семи исследований случай-контроль (2815 случаев, 3170 контролей), в которых конкретно оценивался риск, связанный с курением сигарет по генотипу GSTM1 , показал, что курение было связано с риском только после удаления GSTM1 (30). Хотя общий размер выборки, включенной в эти мета-анализы, дал бы достаточную мощность для обнаружения небольших изменений риска, эти мета-анализы, вероятно, были ошибочными по нескольким причинам. К ним относятся проблемы дизайна (на базе больниц или на уровне населения) и неспособность приспособиться к этнической принадлежности.Что касается этнической принадлежности, традиционно считается, что различия между исследованиями отражают смешение, обусловленное этническим генетическим происхождением или неизмеренными воздействиями, которые различаются между исследуемыми популяциями. С другой стороны, такие различия могут быть связаны (по крайней мере частично) с тем фактом, что генотип с делецией GSTM1 различается по частоте в разных этнических группах (ссылка 20; это исследование) и даже у представителей европеоидной расы (37,5-64,0%). Это может привести к искажению результатов из-за проблем, связанных с дизайном анализа — дизайн анализа на основе геля, который до сих пор использовался почти исключительно для генотипирования этого варианта делеции, препятствовал обнаружению гетерозиготных нулевых носителей отдельно от гомозигот дикого типа.Следовательно, доля гетерозигот в этом пуле будет варьироваться в зависимости от частоты аллелей, как и степень, в которой любой кодоминантный эффект эффективно маскирует риск, связанный с гомозиготной делецией. Интересно отметить, что из метаанализа Sull et al. (20), риск рака груди был значительно увеличен для популяций с более низкой частотой нулевого аллеля GSTM1 (<50,4%), но не для популяций с частотой нулевого аллеля GSTM1 ≥50.4%. Однако, что наиболее важно, в этих предыдущих исследованиях не проводилась систематическая оценка роли внутрипутных взаимодействий, как это делаем мы.
Это первое крупное исследование, изучающее связь между вариантами гена EPHX и GST и риском рака груди. Наши результаты ассоциаций с генотипами GSTT1 и GSTP1 по отдельности согласуются с исследованиями метаанализа, но не с нашими результатами только для генотипа с делецией GSTM1 .Мы также нашли доказательства статистического взаимодействия между генотипами с трехсторонним взаимодействием между генотипами EPHX, GSTT1 и GSTM1 , с оценкой, которая была намного больше, чем для любых отдельных генотипов или других эпидемиологических / демографических переменных, включенных в та же скорректированная модель. Это говорит о том, что риск рака груди в результате таких взаимодействий генов может быть выше, чем для многих известных эпидемиологических факторов риска рака груди. Недавно были представлены аналогичные доказательства вклада в восприимчивость к раку груди двусторонних взаимодействий между полиморфизмами, имеющими вероятную функциональную значимость, в том числе в метаболизме канцерогенов (31).
Хотя существует сильное биологическое обоснование взаимодействий между полиморфизмами в одном и том же пути, мы признаем, что проверили несколько гипотез. Следует проявлять большую осторожность при оценке взаимодействий, особенно тех, которые основаны на небольших размерах клеток, даже если они были получены в результате умеренно большого исследования. Важность очень больших размеров выборки для обеспечения достаточной статистической мощности для достоверного исследования возможных модификаций эффектов или взаимодействий была подчеркнута исследованиями гена метаболизма гормонов CYP17 (32).Таким образом, мы представляем данные таким образом, чтобы другие могли комбинировать их со своими, чтобы попытаться воспроизвести или опровергнуть наши выводы.
Это исследование еще раз подчеркивает необходимость объединения сопоставимых данных из крупных популяционных исследований случай-контроль, чтобы иметь хоть какую-то надежду на установление окончательных взаимодействий ген-ген в предрасположенности к раку груди. В этих исследованиях следует использовать последовательные или сопоставимые меры воздействия, чтобы иметь возможность оценить изменение эффекта от воздействия обычных канцерогенов в окружающей среде, а также скорректировать такие факторы, как социально-экономический статус, в качестве суррогата неизмеренного воздействия (я).Роль семейного реестра больных раком молочной железы (33) и недавно созданного Консорциума ассоциации рака молочной железы (34) будет неотъемлемой частью подтверждения предположительных результатов, подобных тем, о которых сообщается здесь.
Благодарности
Мы благодарим многих женщин, принявших участие в этом исследовании; персонал, проводящий собеседование; врачи, хирурги и онкологи, поддержавшие этот проект; и Р. Минчину из Университета Квинсленда за полезные комментарии к рукописи.
Сноски
↵5 Неопубликованные данные.
Грантовая поддержка: Австралийское семейное исследование рака молочной железы финансировалось Национальным советом по здравоохранению и медицинским исследованиям, Викторианским фондом укрепления здоровья, Советом по борьбе с раком Нового Южного Уэльса, Онкологическим институтом Питера МакКаллума, Мемориальным фондом Инкстер-Росс и Национальным институтом здравоохранения США, в рамках гранта CA 69638, предоставленного Национальным институтом рака молочной железы. Онкологический центр Фокса Чейза, Онкологический институт Хантсмана, Колумбийский университет, Онкологический центр Северной Калифорнии, Онкологический центр Онтарио и Мельбурнский университет.Эта работа финансировалась Национальным советом по здравоохранению и медицинским исследованиям. А.Б. Spurdle финансируется Премией развития карьеры Национального совета по здравоохранению и медицинским исследованиям, G.B. Бирнс финансируется за счет гранта Национального совета по здравоохранению и медицинским исследованиям по наращиванию потенциала в области здоровья населения, а Дж. Ченевикс-Тренч и Дж. Л. Хоппер являются главными и старшими научными сотрудниками Национального совета по здравоохранению и медицинским исследованиям соответственно.
Расходы на публикацию этой статьи были частично покрыты за счет оплаты страницы.Таким образом, данная статья должна быть помечена как реклама в соответствии с 18 U.S.C. Раздел 1734 исключительно для указания этого факта.
Примечание: Дж. Л. Хоппер — руководитель группы консорциума по исследованию рака молочной железы штата Виктория. Содержание этой статьи не обязательно отражает взгляды или политику Национального института рака или любого из сотрудничающих центров в Реестре онкологических семей, а также упоминание торговых наименований, коммерческих продуктов или организаций не подразумевает одобрения U.S. Правительство или Центры регистрации раковых семей.
- Принято 29 января 2007 г.
- Получено 14 сентября 2006 г.
- Исправление получено 17 января 2007 г.
Ссылки
- ↵
Hopper JL, Southey MC, Dite GS, et al. Популяционная оценка среднего возрастного кумулятивного риска рака груди для определенного набора мутаций, усекающих белок в BRCA1 и BRCA2.Эпидемиологические биомаркеры рака до 1999 г .; 8: 741–7.
- ↵
Истон Д., Пето Дж. Вклад наследственной предрасположенности в заболеваемость раком. Cancer Surv 1990; 9: 395–416.
- ↵
Реббек Т.Р., Мартинес М.Э., Селлерс Т.А. и др. Генетическая изменчивость и рак: улучшение условий для публикации ассоциативных исследований. Эпидемиологические биомаркеры рака до 2004 г .; 13: 1985–6.
- ↵
Бирнс Г., Гуррин Л., Даути Дж. И др.Политика публикации или предвзятость публикации? Эпидемиологические биомаркеры рака до 2005 г .; 14: 1363.
- ↵
Oesch F. Эпоксидные гидразы млекопитающих: индуцируемые ферменты, катализирующие инактивацию канцерогенных и цитотоксических метаболитов, полученных из ароматических и олефиновых соединений. Xenobiotica 1973; 3: 305–40.
- ↵
Аданг А.Е., Мейер Д.Д., Брюсси Дж. И др. Взаимодействие крысиных глутатион S -трансфераз 7-7 и 8-8 с γ-глутамил- или глицил-модифицированными аналогами глутатиона.Biochem J 1989; 264: 759–64.
- ↵
Hengstler JG, Arand M, Herrero ME, et al. Полиморфизмы N -ацетилтрансфераз, глутатион-S-трансфераз, микросомальных эпоксидгидролаз и сульфотрансфераз: влияние на предрасположенность к раку. Недавние результаты Cancer Res 1998; 154: 47–85.
- ↵
Странный RC, Фрайер AA. Глутатион S -трансферазы: влияние полиморфизма на предрасположенность к раку.В: Vineis P, редактор. Метаболических полиморфизмов и предрасположенности к раку нет. 148. Лион: Научные публикации МАИР; 1999. стр. 231–49.
- ↵
Hassett C, Aicher L., Sidhu JS, et al. Эпоксидгидролаза микросом человека: генетический полиморфизм и функциональная экспрессия in vitro вариантов аминокислот. Hum Mol Genet 1994; 3: 421–8. Ошибка в: Hum Mol Genet 1994; 3: 1214.
- ↵
Pemble S, Schroeder KR, Spencer SR, et al.Человеческая глутатион S -трансфераза 𝛉 (GSTT1): клонирование кДНК и характеристика генетического полиморфизма. Biochem J 1994; 300: 271–6.
Доска PG. Биохимическая генетика глутатион- S -трансферазы у человека. Am J Hum Genet 1981; 33: 36–43.
- ↵
Seidegard J, Vorachek WR, Pero RW и др. Наследственные различия в экспрессии человеческой глутатионтрансферазы, активной на транс -стильбеноксиде, обусловлены делецией гена.Proc Natl Acad Sci U S. A 1988; 85: 7293–7.
- ↵
Реббек TR. Молекулярная эпидемиология генотипов GSTM1 и GSTT1 глутатиона S -трансферазы человека в отношении предрасположенности к раку. Эпидемиологические биомаркеры рака до 1997 г .; 6: 733–43.
- ↵
Ахмад Х., Уилсон Д.Е., Фриц Р.Р. и др. Первичный и вторичный структурный анализ глутатиона S -трансферазы π из плаценты человека.Arch Biochem Biophys 1990; 278: 398–408.
- ↵
Харрис Л.В., Стаббинс М.Дж., Форман Д. и др. Идентификация генетических полиморфизмов в локусе π глутатиона S -трансферазы и ассоциация с предрасположенностью к раку мочевого пузыря, яичек и простаты. Канцерогенез 1997; 18: 641–4.
- ↵
Зимняк П., Нандури Б., Пикула С. и др. Встречающиеся в природе изоформы глутатиона S -трансферазы GSTP1-1 человека с изолейцином и валином в положении 104 различаются по ферментативным свойствам.Eur J Biochem 1994; 224: 893–9.
Али Осман Ф., Аканде О, Антоун Г. и др. Молекулярное клонирование, характеристика и экспрессия в Escherichia coli полноразмерных кДНК трех вариантов гена π глутатиона человека S -трансферазы. Доказательства дифференциальной каталитической активности кодируемых белков. J Biol Chem 1997; 272: 10004–12.
- ↵
Johansson AS, Stenberg G, Widersten M, et al.Взаимосвязь структура-активность и термостабильность человеческой глутатионтрансферазы P1-1, регулируемая остатком 105 H-сайта. J Mol Biol 1998; 278: 687–98.
- ↵
Egan KM, Cai Q, Shu XO, et al. Генетический полиморфизм GSTM1, GSTP1 и GSTT1 и риск рака груди: результаты Шанхайского исследования рака груди и метаанализа. Эпидемиологические биомаркеры рака до 2004 г .; 13: 197–204.
- ↵
Салл Дж. У., Орр Х, Канг Д. Р. и др.Статус глутатиона S -трансферазы M1 и риск рака груди: метаанализ. Yonsei Med J 2004; 45: 683–9.
- ↵
Vogl FD, Taioli E, Maugard C и др. Глутатион S -трансферазы M1, T1 и P1 и рак груди: объединенный анализ. Эпидемиологические биомаркеры рака до 2004 г .; 13: 1473–9.
- ↵
Сарманова Ю., Сусова С., Gut I, et al. Рак груди: роль полиморфизмов в ферментах биотрансформации.Eur J Hum Genet 2004; 12: 848–54.
- ↵
Hopper JL, Chenevix-Trench G, Jolley DJ, et al. Вопросы дизайна и анализа в популяционном исследовании «случай-контроль-семья» генетической эпидемиологии рака груди и Совместном семейном реестре исследований рака молочной железы (CFRBCS). J Natl Cancer Inst Monogr 1999; 26: 95–100.
- ↵
Dite GS, Jenkins MA, Southey MC, et al. Семейные риски, рак груди с ранним началом и мутации зародышевой линии BRCA1 и BRCA2.J Natl Cancer Inst 2003; 95: 448–57.
- ↵
McCredie MR, Dite GS, Giles GG, et al. Рак груди у австралийских женщин в возрасте до 40 лет. Контроль причин рака, 1998, 9: 189–98.
- ↵
Spurdle AB, Purdie DM, Webb PM и др. Полиморфизм микросомальной эпоксидгидролазы Tyr113His: связь с риском рака яичников. Mol Carcinog 2001; 30: 71–8.
- ↵
Spurdle AB, Webb PM, Purdie DM, et al.Полиморфизмы локусов глутатиона S -трансферазы GSTM1, GSTT1 и GSTP1: риск рака яичников по гистологическим подтипам. Канцерогенез 2001; 22: 67–72.
- ↵
Клейтон Д., Хиллс М. Статистические модели в эпидемиологии. Оксфорд; Нью-Йорк: издательство Оксфордского университета; 1993.
- ↵
Агрести А. Категориальный анализ данных. Нью-Йорк; Брисбен: Уайли; 1990.
- ↵
Терри П.Д., Гудман М.Изменена ли связь между курением сигарет и раком груди из-за генотипа? Обзор эпидемиологических исследований и метаанализа. Эпидемиологические биомаркеры рака до 2006 г .; 15: 602–11.
- ↵
Onay VU, Briollais L, Knight JA, et al. SNP-SNP взаимодействия при предрасположенности к раку груди. BMC Рак 2006; 6: 114.
- ↵
Литтл Дж., Симард Дж. CYP17 и рак груди: нет общего эффекта, но как насчет взаимодействия? Рак молочной железы Res 2005; 7: 238–42.
- ↵
Джон Э.М., Хоппер Дж. Л., Бек Дж. С. и др. Семейный реестр рака груди: инфраструктура для совместных многонациональных, междисциплинарных и трансляционных исследований генетической эпидемиологии рака груди. Рак молочной железы Res 2004; 6: R375–89.
- ↵
Консорциум ассоциации рака молочной железы. Обычно изучаемые однонуклеотидные полиморфизмы и рак груди: результаты Консорциума ассоциации рака груди.J Natl Cancer Inst 2006; 98: 1382–96.
Первичное повреждение ДНК и генетический полиморфизм CYP1A1, EPHX и GSTM1 у рабочих завода по производству графитовых электродов | BMC Public Health
Боффетта П., Журенкова Н., Густавссон П.: Риск рака в результате воздействия полициклических ароматических углеводородов на рабочем месте и в окружающей среде. Контроль причин рака. 1997, 8 (3): 444-472. 10.1023 / А: 1018465507029.
CAS Статья PubMed Google Scholar
Moulin JJ, Wild P, Mur JM, Lafontaine M, Lefer M, Mercier-Gallay M, Villemot P, Whebi V, Coulon JP: Риск рака легких, гортани, глотки и полости рта у рабочих, производящих угольные электроды. Scand J Work Environ Health. 1989, 15 (1): 30-37.
CAS Статья PubMed Google Scholar
Dell’Omo M, Muzi G, Marchionna G, Latini L, Carrieri P, Paolemili P, Abbritti G: Профилактические меры снижают воздействие полициклических ароматических углеводородов на установке графитовых электродов.Occup Environ Med. 1998, 55 (6): 401-406.
Артикул PubMed PubMed Central Google Scholar
Монарка С., Паскини Р., Сфорцолини Г.С., Виола В., Фаджиоли Ф .: Применение анализа мутагенности сальмонелл и определение полициклических ароматических углеводородов на рабочих местах, подверженных воздействию нефтяного пека и нефтяного кокса. Int Arch Occup Environ Health. 1982, 49 (3-4): 223-239. 10.1007 / BF00377932.
CAS Статья PubMed Google Scholar
Паскуини Р., Монарка С., Сфорцолини Г.С., Конти Р., Фаджиоли Ф .: Мутагены в моче рабочих с угольным электродом. Int Arch Occup Environ Health. 1982, 50 (4): 387-395. 10.1007 / BF00377835.
CAS Статья PubMed Google Scholar
Петри Т., Шмид П., Шлаттер С. Воздействие полициклических ароматических углеводородов (ПАУ) в воздухе и экскреция с мочой 1-гидроксипирена рабочими угольных анодных заводов. Ann Occup Hyg. 1996, 40 (3): 345-357.
CAS Статья PubMed Google Scholar
Тета М.Дж., Отт М.Г., Шнаттер А.Р.: Наблюдение за смертностью населения на предприятиях по производству углеродных продуктов. Br JInd Med. 1987, 44 (5): 344-350.
CAS Google Scholar
Мори I: Смертность от рака среди рабочих, производящих искусственные графитовые электроды: результаты 38-летнего наблюдения. Occup Environ Med.2002, 59 (7): 473-480. 10.1136 / oem.59.7.473.
CAS Статья PubMed PubMed Central Google Scholar
Густавссон П., Белландер Т., Йоханссон Л., Салмонссон С.: Наблюдение за смертностью и заболеваемостью раком среди шведских рабочих с графитовыми электродами. Environ Res. 1995, 70 (1): 7-10. 10.1006 / enrs.1995.1039.
CAS Статья PubMed Google Scholar
Донато Ф., Монарка С., Маркиона Г., Росси А., Чичони С., Кьеза Р., Колин Д., Боффетта П.: Смертность от рака и хронических респираторных заболеваний среди рабочих, производящих угольные электроды. Occup Environ Med. 2000, 57 (7): 484-487. 10.1136 / oem.57.7.484.
CAS Статья PubMed PubMed Central Google Scholar
МАИР: Полиядерные ароматические соединения, Часть 1, Химические, экологические и экспериментальные данные.Монография МАИР по оценке канцерогенного риска химических веществ для человека. 1983, Лион (Франция): Международное агентство по изучению рака, 32:
Google Scholar
Crofts F, Taioli E, Trachman J, Cosma GN, Currie D, Toniolo P, Garte SJ: Функциональное значение различных генотипов CYP1A1 человека. Канцерогенез. 1994, 15 (12): 2961-2963. 10.1093 / carcin / 15.12.2961.
CAS Статья PubMed Google Scholar
Коулз Б., Кеттерер Б. Роль глутатиона и трансфераз глутатиона в химическом канцерогенезе. Crit Rev BiochemMol Biol. 1990, 25 (1): 47-70.
CAS Статья Google Scholar
Ингельман-Сундберг М.: Генетическая изменчивость восприимчивости и реакции на токсиканты. Toxicol Lett. 2001, 120 (1-3): 259-268. 10.1016 / S0378-4274 (01) 00278-8.
CAS Статья PubMed Google Scholar
Паванелло С., Клонферо Э .: Биологические индикаторы генотоксического риска и метаболических полиморфизмов. Mutat Res. 2000, 463 (3): 285-308. 10.1016 / S1383-5742 (00) 00051-X.
CAS Статья PubMed Google Scholar
Schoket B, Papp G, Levay K, Mrackova G, Kadlubar FF, Vincze I: Влияние метаболических генотипов на уровни биомаркеров генотоксического воздействия. Mutat Res. 2001, 482 (1-2): 57-69.
CAS Статья PubMed Google Scholar
Bartsch H, Nair U, Risch A, Rojas M, Wikman H, Alexandrov K: генетический полиморфизм генов CYP, отдельно или в комбинации, как модификатор риска рака, связанного с табаком. Биомаркеры эпидемиологии рака Пред. 2000, 9 (1): 3-28.
CAS PubMed Google Scholar
Pastorelli R, Guanci M, Cerri A, Negri E, La Vecchia C, Fumagalli F, Mezzetti M, Cappelli R, Panigalli T, Fanelli R и др .: Влияние наследственных полиморфизмов на глутатион S-трансферазу M1 , микросомальная эпоксидгидролаза, ферменты цитохрома P450 на ДНК и белковые аддукты бензо (а) пирендиолепоксида.Биомаркеры эпидемиологии рака Пред. 1998, 7 (8): 703-709.
CAS PubMed Google Scholar
Рохас М., Александров К., Каскорби И., Брокмоллер Дж., Лихачев А., Пожарисский К., Бувье Г., Обуртин Г., Майер Л., Копп-Шнайдер А. и др.: Диол-эпоксид с высоким содержанием бензо [а] пирена Уровни аддуктов ДНК в легких и клетках крови людей с комбинированными генотипами CYP1A1 MspI / Msp-GSTM1 * 0 / * 0. Фармакогенетика. 1998, 8 (2): 109-118. 10.1097 / 00008571-199804000-00003.
CAS Статья PubMed Google Scholar
Тейшейра Дж. П., Гаспар Дж., Мартиньо Дж., Сильва С., Родригес С., Майан О., Мартин Э., Фермер П. Б., Рюфф Дж.: Уровни аддуктов ароматической ДНК у рабочих коксовой печи: корреляция с полиморфизмами генов GSTP1, GSTM1 , GSTT1 и CYP1A1. Mutat Res. 2002, 517 (1-2): 147-155.
CAS Статья PubMed Google Scholar
Au WW, Oh HY, Grady J, Salama SA, Heo MY: Полезность генетической предрасположенности и биомаркеров для оценки риска для здоровья, связанного с окружающей средой. Environ Mol Mutagen. 2001, 37 (3): 215-225. 10.1002 / em.1030.
CAS Статья PubMed Google Scholar
Autrup H: Генетические полиморфизмы в ферментах метаболизма xenobiotica человека как факторы чувствительности при токсической реакции. Mutat Res. 2000, 464 (1): 65-76.
CAS Статья PubMed Google Scholar
Raaka S, Hassett C, Omiencinski CJ: микросомальная эпоксидгидролаза человека: генетический полиморфизм 5′-фланкирующей области. Канцерогенез. 1998, 19 (3): 387-393. 10.1093 / carcin / 19.3.387.
CAS Статья PubMed Google Scholar
Борд П., Когган М., Джонстон П., Росс В., Сузуки Т., Уэбб Г.: Генетическая гетерогенность трансфераз глутатиона человека: комплекс семейств генов. Pharmacol Ther. 1990, 48 (3): 357-369. 10.1016 / 0163-7258 (90)
-6.
CAS Статья PubMed Google Scholar
Бинкова Б., Топинка Дж., Мрацкова Г., Гайдосова Д., Видова П., Ставкова З., Петерка В., Пильчик Т., Римар В., Добиас Л. и др.: Исследование рабочих коксовой печи: влияние воздействия и GSTM1 и генотипы NAT2 по уровням аддукта ДНК в лейкоцитах и лимфоцитах, как определено с помощью 32P-пост-мечения. Mutat Res. 1998, 416 (1-2): 67-84.
CAS Статья PubMed Google Scholar
Alexandrie AK, Warholm M, Carstensen U, Axmon A, Hagmar L, Levin JO, Ostman C, Rannug A: Полиморфизмы CYP1A1 и GSTM1 влияют на уровни 1-гидроксипирена в моче после воздействия ПАУ. Канцерогенез. 2000, 21 (4): 669-676. 10.1093 / carcin / 21.4.669.
CAS Статья PubMed Google Scholar
Срам Р.Дж., Бинкова Б. Исследования молекулярной эпидемиологии воздействия мутагенов и канцерогенов на рабочем месте и в окружающей среде, 1997–1999.Перспектива здоровья окружающей среды. 2000, 108 (Приложение 1): 57-70. 10.2307 / 3454632.
CAS Статья PubMed PubMed Central Google Scholar
Jongeneelen FJ, Anzion RB, Henderson PT: Определение гидроксилированных метаболитов полициклических ароматических углеводородов в моче. J Chromatogr. 1987, 413: 227-232. 10.1016 / 0378-4347 (87) 80230-X.
CAS Статья PubMed Google Scholar
VanRooij JG, Bodelier-Bade MM, Jongeneelen FJ: Оценка индивидуального кожного и респираторного поглощения полициклических ароматических углеводородов у 12 работников коксовых печей. Br J Ind Med. 1993, 50 (7): 623-632.
CAS PubMed PubMed Central Google Scholar
Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A, Kobayashi H, Miyamae Y, Rojas E, Ryu JC, Sasaki YF: Анализ одноклеточного геля / кометы: руководство для in vitro и in vivo генетическое токсикологическое тестирование.Environ Mol Mutagen. 2000, 35 (3): 206-221. 10.1002 / (SICI) 1098-2280 (2000) 35: 3 <206 :: AID-EM8> 3.0.CO; 2-J.
CAS Статья PubMed Google Scholar
Pastorelli R, Cerri A, Rozio M, Dell’Omo M, Muzi G, Abbritti G, Fanelli R, Airoldi L: Аддукты бензо (а) пирендиолепоксида к альбумину у рабочих, подвергшихся воздействию полициклических ароматических углеводородов: ассоциация со специфическими генотипами CYP1A1, GSTM1, GSTP1 и EHPX. Биомаркеры.2001, 6 (5): 357-374. 10.1080 / 13547500110044267.
CAS Статья PubMed Google Scholar
Hayashi S, Watanabe J, Kawajiri K: Генетические полиморфизмы в 5′-фланкирующей области изменяют регуляцию транскрипции гена цитохрома P450IIE1 человека. J Biochem (Токио). 1991, 110 (4): 559-565.
CAS Google Scholar
Cascorbi I, Brockmoller J, Roots I: Полиморфизм C4887A в экзоне 7 человеческого CYP1A1: популяционная частота, мутации и влияние на восприимчивость к раку легких.Cancer Res. 1996, 56 (21): 4965-4969.
CAS PubMed Google Scholar
Bell DA, Taylor JA, Paulson DF, Robertson CN, Mohler JL, Lucier GW: Генетический риск и воздействие канцерогенов: распространенный наследственный дефект гена метаболизма канцерогенов, глутатион-S-трансферазы M1 (GSTM1), который увеличивает предрасположенность к раку мочевого пузыря. J Natl Cancer Inst. 1993, 85 (14): 1159-1164. 10.1093 / jnci / 85.14.1159.
CAS Статья PubMed Google Scholar
Brendler-Schwaab SY, Schmezer P, Liegibel U, Weber S, Michalek K, Tompa A, Pool-Zobel BL: Клетки различных тканей для токсикологических исследований in vitro и in vivo: Сборник методов выделения. Токсикология in vitro. 1994, 8 (6): 1285-1302. 10.1016 / 0887-2333 (94) -8.
CAS Статья PubMed Google Scholar
Jones KH, Senft JA: Улучшенный метод определения жизнеспособности клеток путем одновременного окрашивания флуоресцеина диацетат-пропидий йодид.J Histochem Cytochem. 1985, 33 (1): 77-79.
CAS Статья PubMed Google Scholar
Сингх Н.П., Маккой М.Т., Тайс Р.Р., Шнайдер Е.Л.: простой метод количественного определения низких уровней повреждения ДНК в отдельных клетках. Exp Cell Res. 1988, 175 (1): 184-191. 10.1016 / 0014-4827 (88)-0.
CAS Статья PubMed Google Scholar
Villarini M, Moretti M, Pasquini R, Scassellati-Sforzolini G, Fatigoni C, Marcarelli M, Monarca S, Rodriguez AV: Генотоксические эффекты инсектицида дельтаметрина in vitro на лейкоциты периферической крови человека: повреждение ДНК (‘ комета) в отношении индукции обменов сестринских хроматид и микроядер.Токсикология. 1998, 130 (2-3): 129-139. 10.1016 / S0300-483X (98) 00097-3.
CAS Статья PubMed Google Scholar
Villarini M, Scassellati-Sforzolini G, Moretti M, Pasquini R: Генотоксичность тербутрина in vitro, оцененная с помощью щелочного одноклеточного микрогелевого электрофореза «кометный» анализ. Cell Biol Toxicol. 2000, 16 (5): 285-292. 10.1023 / А: 10267
308.
CAS Статья PubMed Google Scholar
Moretti M, Villarini M, Scassellati-Sforzolini G, Monarca S, Salucci A, Vicent Rodriguez A: Применение одноклеточного гель-электрофореза («комета») для обнаружения первичных повреждений ДНК у рабочих резиновой промышленности : сравнение ручного и компьютерного анализа. Токсикологическая и экологическая химия. 1999, 13-24.
Google Scholar
Bonassi S, Fontana V, Ceppi M, Barale R, Biggeri A: Анализ коррелированных данных в исследованиях биомониторинга человека.Случай клеток с высокой частотой обмена сестринских хроматид. Mutat Res. 1999, 438 (1): 13-21.
CAS Статья PubMed Google Scholar
Коллинз А.Р.: Кометный анализ повреждений и репарации ДНК: принципы, применения и ограничения. Mol Biotechnol. 2004, 26 (3): 249-261. 10.1385 / МБ: 26: 3: 249.
CAS Статья PubMed Google Scholar
Nan HM, Kim H, Lim HS, Choi JK, Kawamoto T., Kang JW, Lee CH, Kim YD, Kwon EH: Влияние занятий, образа жизни и генетических полиморфизмов CYP1A1, CYP2E1, GSTM1 и GSTT1 на 1-гидроксипирен в моче и Концентрации 2-нафтола. Канцерогенез. 2001, 22 (5): 787-793. 10.1093 / carcin / 22.5.787.
CAS Статья PubMed Google Scholar
Ovrebo S, Haugen A, Fjeldstad PE, Hemminki K, Szyfter K: Биологический мониторинг воздействия полициклических ароматических углеводородов на заводе по производству электродной пасты.J Occup Med. 1994, 36 (3): 303-310. 10.1097 / 00043764-1900-00007.
CAS Статья PubMed Google Scholar
Siwinska E, Mielzynska D, Kapka L: Связь между 1-гидроксипиреном в моче и генотоксическими эффектами у работников коксовых печей. Occup Environ Med. 2004, 61 (3): e10-10.1136 / oem.2002.006643.
CAS Статья PubMed PubMed Central Google Scholar
Duez P, Dehon G, Kumps A, Dubois J: Статистика анализа комет: ключ к различению генотоксических эффектов. Мутагенез. 2003, 18 (2): 159-166. 10.1093 / mutage / 18.2.159.
CAS Статья PubMed Google Scholar
Buschini A, De Palma G, Poli P, Martino A, Rossi C, Mozzoni P, Scotti E, Buzio L, Bergamaschi E, Mutti A: Генетический полиморфизм ферментов метаболизма лекарств и индуцированное стиролом повреждение ДНК .Environ Mol Mutagen. 2003, 41 (4): 243-252. 10.1002 / em.10150.
CAS Статья PubMed Google Scholar
Лу И, Моримото К., Накаяма К.: Практика здоровья и повреждение ДНК лейкоцитов у японских рабочих, занимающихся твердосплавными работами. Предыдущая Мед. 2006, 43 (2): 140-144. 10.1016 / j.ypmed.2006.03.008.
CAS Статья PubMed Google Scholar
Sigurdson AJ, Hauptmann M, Alexander BH, Doody MM, Thomas CB, Struewing JP, Jones IM: повреждение ДНК при раке щитовидной железы и множественные случаи рака, контрольные и долгоживущие люди.Mutat Res. 2005, 586 (2): 173-188.
CAS Статья PubMed Google Scholar
Carstensen U, Hou SM, Alexandrie AK, Hogstedt B, Tagesson C, Warholm M, Rannug A, Lambert B, Axmon A, Hagmar L: Влияние генетического полиморфизма ферментов биотрансформации на мутации генов, разрывы цепей дезоксирибонуклеиновая кислота, микроядра в мононуклеарных клетках крови и 8-гидроксидезоксигуанозин в моче у рабочих, подвергшихся воздействию полиароматических углеводородов.Scand J Work Environ Health. 1999, 25 (4): 351-360.
CAS Статья PubMed Google Scholar
Crebelli R, Carta P, Andreoli C, Aru G, Dobrowolny G, Rossi S, Zijno A: Биомониторинг работников первичной алюминиевой промышленности: обнаружение микроядер и восстанавливаемые повреждения ДНК с помощью щелочного SCGE. Mutat Res. 2002, 516 (1-2): 63-70.
CAS Статья PubMed Google Scholar
Marczynski B, Rihs HP, Rossbach B, Holzer J, Angerer J, Scherenberg M, Hoffmann G, Bruning T., Wilhelm M: анализ 8-оксо-7,8-дигидро-2′-дезоксигуанозина и разрывов цепей ДНК белого цвета клетки крови рабочих, подвергшихся профессиональному облучению: сравнение с мониторингом окружающей среды, метаболиты в моче и полиморфизм ферментов. Канцерогенез. 2002, 23 (2): 273-281. 10.1093 / carcin / 23.2.273.
CAS Статья PubMed Google Scholar
Moretti M, Villarini M, Scassellati-Sforzolini G, Monarca S, Libraro M, Fatigoni C, Donato F, Leonardis C, Perego L: Биологический мониторинг генотоксической опасности у рабочих резиновой промышленности. Перспектива здоровья окружающей среды. 1996, 104 (Дополнение 3): 543-545. 10.2307 / 3432820.
CAS Статья PubMed PubMed Central Google Scholar
Sul D, Oh E, Im H, Yang M, Kim CW, Lee E: Повреждение ДНК в T- и B-лимфоцитах и гранулоцитах у рабочих, занимающихся инспекцией выбросов и сжиганием, подвергшихся воздействию полициклических ароматических углеводородов.Mutat Res. 2003, 538 (1-2): 109-119.
CAS Статья PubMed Google Scholar
van Delft JH, Steenwinkel MS, van Asten JG, de Vogel N, Bruijntjes-Rozier TC, Schouten T., Cramers P, Maas L, van Herwijnen MH, van Schooten F, et al: Биологический мониторинг воздействия к полициклическим ароматическим углеводородам коксоводов в связи с курением и генетическим полиморфизмом для GSTM1 и GSTT1. Ann Occup Hyg. 2001, 45 (5): 395-408.
CAS Статья PubMed Google Scholar
Hoffmann H, Hogel J, Speit G: Влияние курения на эффекты ДНК в анализе комет: метаанализ. Мутагенез. 2005, 20 (6): 455-466. 10.1093 / mutage / gei064.
CAS Статья PubMed Google Scholar
Cosma G, Crofts F, Currie D, Wirgin I, Toniolo P, Garte SJ: Расовые различия в полиморфизмах длины рестрикционных фрагментов и индуцируемости матричной РНК гена CYP1A1 человека.Биомаркеры эпидемиологии рака Пред. 1993, 2 (1): 53-57.
CAS PubMed Google Scholar
Чжао Х., Шпиц М.Р., Гвин К.М., Ву Х: Микросомальные полиморфизмы эпоксидгидролазы и риск рака легких у неиспаноязычных белых. Mol Carcinog. 2002, 33 (2): 99-104. 10.1002 / mc.10023.
CAS Статья PubMed Google Scholar
Hirvonen A, Husgafvel-Pursiainen K, Anttila S, Vainio H: нулевой генотип GSTM1 как модификатор потенциального риска плоскоклеточного рака легкого.Канцерогенез. 1993, 14 (7): 1479-1481. 10.1093 / carcin / 14.7.1479.
CAS Статья PubMed Google Scholar
Somorovska M, Jahnova E, Tulinska J, Zamecnikova M, Sarmanova J, Terenova A, Vodickova L, Liskova A, Vallova B, Soucek P, et al: Биомониторинг профессионального воздействия стирола на установке для ламинирования пластмасс . Mutat Res. 1999, 428 (1-2): 255-269.
CAS Статья PubMed Google Scholar
Кульюкка-Рабб Т., Нюлунд Л., Вааранринта Р., Савела К., Мутанен П., Вейдебаум Т., Сорса М., Раннуг А., Пелтонен К.: Влияние соответствующих генотипов на биомаркеры, связанные с воздействием ПАУ. J Expo Anal Environ Epidemiol. 2002, 12 (1): 81-91. 10.1038 / sj.jea.7500204.
CAS Статья PubMed Google Scholar
Апостоли П., Нери Дж., Лукас Д., Манно М., Берту Ф .: Влияние генетических полиморфизмов CYP1A1 и GSTM1 на уровни 1-гидроксипирена в моче.Toxicol Lett. 2003, 144 (1): 27-34. 10.1016 / S0378-4274 (03) 00227-3.
CAS Статья PubMed Google Scholar
Pavanello S, Pulliero A, Siwinska E, Mielzynska D, Clonfero E: Сниженная эксцизионная репарация нуклеотидов и GSTM1-нулевые генотипы влияют на уровни аддукта анти-B [a] PDE-ДНК в мононуклеарных белых кровяных тельцах с высоким содержанием PAH -открытые работники коксовых печей. Канцерогенез. 2005, 26 (1): 169-175. 10.1093 / carcin / bgh403.
CAS Статья PubMed Google Scholar
Рохас М., Каскорби И., Александров К., Крик Э., Обуртин Г., Майер Л., Копп-Шнайдер А., Рутс I, Барч Н.: Модуляция уровней аддукта бензо [а] пирендиолепоксида-ДНК в лейкоцитах человека с помощью CYP1A1, Полиморфизм GSTM1 и GSTT1. Канцерогенез. 2000, 21 (1): 35-41. 10.1093 / carcin / 21.1.35.
CAS Статья PubMed Google Scholar
Cotelle S, Ferard JF: Анализ комет в генетической экотоксикологии: обзор. Environ Mol Mutagen.1999, 34 (4): 246-255. 10.1002 / (SICI) 1098-2280 (1999) 34: 4 <246 :: AID-EM4> 3.0.CO; 2-V.
CAS Статья PubMed Google Scholar
Merlo DF, Garattini S, Gelatti U, Simonati C, Covolo L, Ceppi M, Donato F: когортное исследование смертности среди рабочих завода по производству графитовых электродов в Италии. Occup Environ Med. 2004, 61 (2): e9-10.1136 / oem.2003.009357.
CAS Статья PubMed PubMed Central Google Scholar
Полиморфизм в гене микросомальной эпоксидгидролазы связан с преэклампсией
Преэклампсия представляет собой одно из наиболее серьезных распространенных медицинских осложнений беременности у человека.Однако факторы, стимулирующие начало и прогрессирование болезненного процесса, остаются неуловимыми. Появляется все больше доказательств того, что дисбаланс между токсичными соединениями, такими как перекиси липидов и свободные радикалы кислорода, и веществами, выводящими токсины и поглощающими вещества, может вносить вклад в патофизиологию преэклампсии.
Многие ферменты, включая цитохромы P450, глутатион S-трансферазы, 5 и эпоксидгидролазы, метаболизируют эндогенные и экзогенные соединения. 6-8 Микросомальная эпоксидгидролаза (EPHX, EC 3.3.2.3) катализирует гидролиз оксидов арена и щелочей с образованием транс- -дигидродиолов.9 10 Хотя этот гидролиз обычно приводит к детоксикации, поскольку он дает меньше реакционноспособных и более водорастворимых соединений, некоторые производные дигидродиола, особенно в сочетании с окислительным метаболизмом цитохрома P450, являются субстратами для дополнительного метаболизма, в результате чего образуются более реактивные и мутагенные соединения, которые могут связываться с геномной ДНК. .6 8 11 Генетический полиморфизм был описан в гене человека, кодирующем микросомальную эпоксидгидролазу ( EPHX ).Два из этих полиморфизмов, 113Tyr → His в экзоне 3 и 139His → Arg в экзоне 4, были связаны со снижением или увеличением активности фермента соответственно.12 Вариации активности фермента для EPHX в результате таких полиморфизмов могут приводить к изменению индивидуальная предрасположенность к таким заболеваниям, как преэклампсия и HELLP-синдром. Однако пока неизвестно, какие реактивные промежуточные соединения, образованные EPHX, могут быть биологически вероятными с точки зрения предрасположенности к преэклампсии. В этом исследовании случай-контроль мы исследовали возможную связь между генотипами EPHX и риском преэклампсии и HELLP-синдрома.Кроме того, три предполагаемых фенотипа активности фермента EPHX в результате сочетания обоих полиморфизмов были оценены в соответствии с исследованием Hassett et al 12 функциональной экспрессии вариантных аллелей in vitro по остатку 113 (экзон 3, медленный аллель His113) и по остатку 139 (экзон 4, быстрый аллель Arg139). Субъекты с низкой активностью были гомозиготными по His113 или гетерозиготными по His113 в сочетании с гомозиготностью по His139. Субъекты с промежуточной активностью были гомозиготными по Tyr113 и His139 или гетерозиготными по Tyr113 и His139.Субъекты с высокой активностью были гомозиготными по Arg139 или гетерозиготными по Arg139 в сочетании с гомозиготностью по Tyr113.
Материалы и методы
ПАЦИЕНТОВ
Исследовательская группа состояла из 181 белой голландской женщины с преэклампсией в анамнезе с (n = 96) или без (n = 87) синдромом HELLP, которые были описаны ранее.13 Наблюдательный совет учреждения одобрил экспериментальный протокол. Преэклампсия определялась как возникновение после 20 недель беременности диастолического артериального давления выше 90 мм рт. Ст. (Фаза 5 звук Короткова) в двух или более случаях с интервалом не менее четырех часов с протеинурией (уровень белка в моче более 0.3 г / л за 24-часовой период сбора мочи). Синдром HELLP был определен как одновременное возникновение количества тромбоцитов менее 100 × 10 9 / л, концентрации аспартатаминотрансферазы и аланинаминотрансферазы в сыворотке выше 30 МЕ / л, а также гемолиза, установленного на основании аномального мазка крови. Группа из 151 случайно выбранной небеременной белой голландской женщины без основного диабета или почечного, печеночного, гематологического, ранее существовавшего гипертонического или сердечно-сосудистого заболевания, у которых была только нормальная нормальная беременность (среднее количество беременностей 2, диапазон от 1 до 4 ) служил контролем.Общие популяционные характеристики контрольных групп и случаев в период, когда они были госпитализированы по поводу преэклампсии, приведены в таблице 1.
Таблица 1Характеристики пациентов и контрольных групп в период госпитализации по поводу преэклампсии
АНАЛИЗ МУТАЦИЙ
После получения информированного согласия был проведен забор крови. Кровь собирали венепункцией в стерильные, силиконизированные, вакуумные пробирки с этилендиаминтетрауксусной кислотой (ЭДТА) объемом 4 мл (Becton Dickinson, Meylan cedex, Франция).Сразу после сбора кровь хранили при -20 ° C до анализа. Геномную ДНК выделяли из цельной крови с использованием набора для очистки геномной ДНК Wizard TM в соответствии с инструкциями производителя (Promega, Мэдисон, Висконсин, США).
Были исследованы два отдельных полиморфизма в EPHX , а именно 113Tyr → His в экзоне 3 и 139His → 39Arg в экзоне 4. Сначала образцы ДНК амплифицировали с помощью полимеразной цепной реакции (ПЦР) с использованием модификации метода, описанного Hassett et al .12 Реакционная смесь ПЦР содержала примерно 100 нг геномной ДНК, 50 ммоль / л KCl, 10 ммоль / л Tris HCl, pH 8,0, 2,0 ммоль / л MgCl 2 , 1,5 ед. Полимеразы Taq и 1 мкмоль. / л каждого из конкретных праймеров прямой (смысловой) и обратной (антисмысловой) амплификации в общем объеме 50 мкл. ДНК денатурировали при 94 ° C в течение пяти минут. Тридцать циклов амплификации начинались с денатурации при 93 ° C, отжига праймеров при 58 ° C для экзона 3 и 65 ° C для экзона 4 и удлинения при 72 ° C, каждый этап длился одну минуту.Для обнаружения варианта 113Tyr → His продукт ПЦР расщепляли Tth 111 и затем подвергали электрофорезу в 3% агарозном геле. Аллель 113Tyr был идентифицирован по непереваренной единственной полосе ДНК из 231 пары оснований (п.н.), тогда как кодирующий аллель 113His дал две полосы ДНК по 209 п.н. и 22 п.н. Для обнаружения варианта 139His → Arg продукт ПЦР расщепляли Rsa I и затем прогоняли в 3% агарозном геле. Аллель, кодирующий 139His, был идентифицирован двумя полосами ДНК (295 и 62 п.н.), тогда как аллель 139Arg давал три полосы ДНК после переваривания (174, 121 и 62 п.н.).Компания Pharmacia Biotech (Розендал, Нидерланды) синтезировала все праймеры, и все химические вещества, необходимые для ПЦР, были приобретены у Promega (Мэдисон, Висконсин, США).
СТАТИСТИЧЕСКИЙ АНАЛИЗ
Клинические характеристики между группами сравнивали с использованием U-критерия Манна-Уитни. Статистическая значимость различий для индивидуальных полиморфизмов между разными группами и прогнозируемой активности фермента (высокая, средняя или низкая активность) была проверена с помощью анализа хи-квадрат (χ 2 ) с поправкой Йетса в таблицах сопряженности 2 × 3.Статистическая значимость была принята как p <0,05. Все статистические анализы были выполнены с помощью пакета статистических программ SPSS (SPSS Inc, Чикаго, США).
Результаты
Средний возраст между случаями и контрольной группой существенно не отличался. Не было значительных различий в сроке беременности, рождении, материнском возрасте или диастолическом артериальном давлении между преэкламптическими группами с или без HELLP-синдрома.
Распределение полиморфных вариантов в экзоне 3 и экзоне 4 гена EPHX у пациентов и контрольной группы суммировано в таблице 2.
Таблица 2Распределение полиморфных вариантов в экзонах 3 и 4 гена EPHX
В контрольной группе кодирующий аллель 113His в экзоне 3, соответствующий низкой активности фермента, встречался чаще, чем кодирующий аллель 113Tyr. Шестьдесят семь (44%) контролей были гомозиготными по полиморфизму экзона 3 (His113 His113) и 60 (40%) были гетерозиготными (Tyr113 His113). Мы обнаружили статистически значимо более высокую частоту быстрого генотипа Tyr113 Tyr113 в экзоне 3 среди женщин с преэклампсией в анамнезе (29%) по сравнению с контрольной группой (16%, отношение шансов (OR) 2.0, 95% доверительный интервал (ДИ) 1,2–3,7) (χ 2 = 9,99, p = 0,0068) при сравнении генотипа Tyr113 Tyr113 с генотипами Tyr113 His113 и His113 His113. Подгруппа женщин с преэклампсией с HELLP в анамнезе по сравнению с группой преэклампсии без HELLP не показала значимых различий в встречаемости полиморфных вариантов в экзоне 3 (OR 1,4, 95% CI 0,7-2,8) ( χ 2 = 0,83, p = 0,66). Не было значительных различий в показателях полиморфизма экзона 3 между женщинами с преэклампсией с (n = 98) или без (n = 85) задержкой внутриутробного развития плода (ЗВУР, определяемой как масса тела при рождении ниже 10-го центиля).Двадцать девять процентов случаев с IUGR имели генотип Tyr113 Tyr113 по сравнению с 22% случаев без IUGR (OR 1,2, 95% CI 0,6–2,4, p = 0,58).
Только 3% контролей были гомозиготными по полиморфизму экзона 4 (Arg139 Arg139), а 33% из 151 субъекта были гетерозиготными (His139 Arg139), по сравнению с 7% и 37%, соответственно, преэкламптических случаев с или без АД-синдром. Это привело к незначительному OR 2,2 (95% ДИ 0,7-7,4, p = 0,20) при сравнении генотипа Arg139 Arg139 с генотипом His139 Arg139 и His139 His139.Опять же, подгруппа женщин с преэклампсией в анамнезе с HELLP, по сравнению с группой без HELLP, не показала значимых различий в частоте полиморфных вариантов в экзоне 4 (OR 0,9, 95% CI 0,3-3,3, p = 0,85).
Частоты аллелей для экзона 3 и экзона 4 не различались между случаями и контролем (χ 2 = 1,90, p = 0,17 для экзона 3; χ 2 = 2,88, p = 0,10 для экзона 4, соответственно).
Распределение субъектов с прогнозируемой высокой, средней и низкой активностью EPHX составило 16 человек.0, 26,9 и 57,1%, соответственно, среди преэкламптических женщин и 4,5%, 18,2% и 77,3%, соответственно, среди контрольной группы. Была обнаружена значительная связь между прогнозируемыми уровнями активности ферментов и преэклампсией (p = 0,0009), что привело к более высокому риску при более высоких прогнозируемых уровнях ферментов.
Обсуждение
Дисбаланс между токсичными и детоксицирующими веществами может играть роль в патофизиологии преэклампсии. Эпоксидгидролаза микросом является одним из ферментов, играющих важную роль в большом количестве процессов детоксикации и в метаболизме эндогенных и экзогенных соединений.В этом исследовании случай-контроль мы попытались выяснить, могут ли вариации в скорости генетического полиморфизма в гене EPHX коррелировать с предрасположенностью к преэклампсии с синдромом HELLP или без него.
Значительная разница в частотах Tyr113 Tyr113 генотипа EPHX наблюдалась между преэкламптической и контрольной группами. Hassett и др. 12 показали, что замена аллеля гистидина 113 на тирозин 113 снижает активность EPHX примерно на 40%, тогда как замена гистидина 139 аргинином 139 увеличивает активность фермента на 25%.Наше исследование показывает, что гомозиготный генотип в экзоне 3 гена EPHX , приводящий к Tyr113 Tyr113 и соответствующий высокой активности фермента, связан с повышенной частотой преэклампсии. Кроме того, прогнозируемый высокий уровень активности EPHX, оцененный по полиморфизму в экзонах 3 и 4 вместе, был связан с преэклампсией. Связь между более высокой частотой генотипа Tyr113 Tyr113 с высокой активностью фермента EPHX в преэкламптической группе и риском преэклампсии может отражать усиленную активацию эндогенными или экзогенными субстратами более реактивных мутагенных производных диола у этих пациентов.
До сих пор было выполнено лишь несколько исследований роли EPHX в репродуктивной системе. Ван и др. 14 недавно показали, что полиморфизм в гене EPHX может влиять на предрасположенность к самопроизвольному аборту. Они показали связь между генотипом His113 His113 и повышенным риском самопроизвольного аборта. Напротив, Lancaster et al 15 обнаружили связь между генотипом Tyr113 и Tyr113 в экзоне 3 и раком яичников.Эти различные результаты могут быть связаны с тем, что в большинстве случаев EPHX играет роль детоксикации, не позволяя высокореактивным эпоксидам модифицировать основные клеточные (макро) молекулы. Однако при определенных условиях EPHX может способствовать активации других соединений, что приводит к отравлению, а не к детоксикации. 11 16 17 Более того, EPHX может участвовать в реакциях стероидогенеза, что может объяснить наблюдение связи между медленным генотипом и защитой от рака яичников.15
Полиморфизмы в гене EPHX также связаны с гепатоцеллюлярным раком, эмфиземой и раком легких.18-20 Эти результаты могут представлять собой пример органоспецифических различий в восприимчивости к заболеванию в результате генетических полиморфизмов, поскольку генотип EPHX с высокой активностью Tyr113 Tyr113 может защищать от самопроизвольного аборта14 и гепатоцеллюлярного рака 18, тогда как восприимчивость к раку яичников 15, раку легких 20 и преэклампсии может повышаться.
Частоты полиморфных вариантов в экзоне 4 гена EPHX не сильно различаются в нескольких исследуемых популяциях.12 14 19 20 Однако сообщалось о заметных вариациях в показателях полиморфизма в экзоне 3 для разных этнических и географических популяций; диапазоны от 26-69% до 22-56% были зарегистрированы для генотипов Tyr113 Tyr113 и Tyr113 His113 соответственно12. 14 15 18-20 Зарегистрированные частоты генетических полиморфизмов в гене EPHX перечислены в таблице 3. Поразительно, что наши данные о полиморфизмах экзона 3 у белых субъектов больше похожи на опубликованные данные в китайской контрольной группе, чем в других белых популяциях. .14 18
Таблица 3Частоты генетических полиморфизмов в гене EPHX в контрольных парах
Эти данные подтверждают гипотезу о том, что генетические вариации генов, кодирующих ферменты, участвующие в метаболизме экзогенных или эндогенных веществ, могут влиять на риск заболеваний беременности, таких как самопроизвольный аборт и преэклампсия.
(PDF) Полиморфизм в гене микросомальной эпоксидгидролазы связан с преэклампсией
(n = 96) или без (n = 87) дрома HELLP-син-
, который был описан ранее.
13
Наблюдательный совет учреждения одобрил экспериментальный протокол
. Преэклампсия была определена как возникновение после 20 недель беременности
диастолического артериального давления выше 90
мм рт.
протеинурия (белок в моче более 0,3 г / л
за 24-часовой период сбора мочи). Синдром
HELLP был определен как одновременное появление количества тромбоцитов менее
100 × 10
9
/ л, сывороточной аспартатаминотрансферазы
и концентрации аланинаминотрансферазы
в сыворотке выше 30 МЕ / л и гемолиз, установленный на основании патологического мазка крови.Группа
из 151 случайно выбранной небеременной женщины,
белых голландских женщин без основного диабета
или почечного, печеночного, гематологического,
ранее существовавшего гипертонического или сердечно-сосудистого заболевания,
легкости, у которых были нормальные нормотен-
только беременных (среднее число беременных
нанок 2, диапазон от 1 до 4) служили в качестве контроля.
Общая популяционная характеристика контрольных групп
и случаев в период, когда они были госпитализированы с преэклампсией
, приведены в таблице 1.
АНАЛИЗ МУТАЦИЙ
После получения информированного согласия был проведен забор крови
. Кровь собирали путем венепункции
в стерильные, силиконизированные, этилен-
диаминтетрауксусной кислоты (ЭДТА) 4 мл вакуумные пробирки
(Becton Dickinson, Meylan cedex,
Франция). Сразу после сбора кровь
хранилась при -20 ° C до анализа. Геномную ДНК
выделяли из цельной крови с использованием набора для очистки геномной ДНК
Wizard
TM
, ac-
в соответствии с инструкциями производителя
(Promega, Мэдисон, Висконсин, США).
Были исследованы два отдельных полиморфизма в EPHX,
, а именно 113Tyr → His в экзоне 3 и
139His → 39Arg в экзоне 4.
Сначала образцы ДНК амплифицировали с помощью полимеразной цепной реакции (ПЦР)
с использованием модификации
метода, описанного Has-
set et al.
12
Реакционная смесь для ПЦР содержала
примерно 100 нг геномной ДНК, 50
ммоль / л KCl, 10 ммоль / л Трис-HCl, pH 8,0, 2.0
ммоль / л MgCl
2
, 1,5UofTa q полимеразы и 1
мкмоль / л каждый из специфических праймеров прямой (смысловой) и
обратной (антисмысловой) амплификации в общем объеме
50 мкл. ДНК денатурировали при
94 ° C в течение пяти минут. Тридцать циклов амплификации катиона
начинались с денатурации при 93 ° C, отжига праймера
при 58 ° C для экзона 3 и 65 ° C для
экзона 4 и удлинения при 72 ° C, каждый шаг для
экзона. минута.Для обнаружения варианта 113Tyr → His
продукт ПЦР расщепляли
Tth211 и затем подвергали электрофорезу
в 3% агарозном геле. Аллель 113Tyr был
, идентифицированный непереваренной одиночной полосой ДНК
из 231 пары оснований (п.н.), тогда как кодирующий аллель 113His
давал две полосы ДНК по 209 п.н.,
,и 22 п.н. Для обнаружения варианта 139His → Arg,
продукт ПЦР расщепляли RsaI и
, затем прогоняли в 3% агарозном геле.Кодирующий аллель 139His
был идентифицирован двумя полосами ДНК
(295 и 62 п.н.), тогда как аллель 139Arg
давал три полосы ДНК после расщепления
(174, 121 и 62 п.н.). Компания Pharmacia Biotech
(Розендал, Нидерланды) синтезировала все праймеры
, и все химические вещества, необходимые для ПЦР
, были приобретены у Promega (Мэдисон, Висконсин,
США).
СТАТИСТИЧЕСКИЙ АНАЛИЗ
Клинические характеристики между группами были
по сравнению с использованием U-критерия Манна-Уитни.
Статистическая значимость различий для
индивидуальных полиморфизмов между различными
группами и прогнозируемая активность фермента
(высокая, средняя или низкая активность) была протестирована
с помощью анализа хи-квадрат (÷
2
). с поправкой Йейтса
в таблицах непредвиденных обстоятельств 2 × 3. Статистическая значимость
была принята за p <0,05. Все статистические анализы
были выполнены с помощью пакета статистических программ SPSS
(SPSS Inc, Chi-
cago, США).
РЕЗУЛЬТАТЫ
Средний возраст между случаями и контрольной группой
не отличался существенно. Не было
значительных различий в сроке беременности, рождении,
материнском возрасте или диастолическом артериальном давлении
между группами преэклампсии с синдромом HELLP или
без него.
Распределение полиморфных вариантов в
как экзоне 3, так и экзоне 4 гена EPHX у
пациентови контрольной группы суммировано в таблице 2.
В контрольной группе 113His, кодирующий аллель
в экзоне 3, соответствующий низкой активности фермента
, встречался чаще, чем кодирующий аллель 113Tyr
. Шестьдесят семь (44%) из контрольных
были гомозиготными по полиморфизму экзона 3
(His113 His113) и 60 (40%) были
гетерозиготными (Tyr113 His113). Мы обнаружили статистически значимо более высокую частоту
быстрого генотипа
Tyr113 Tyr113 в экзоне 3 среди
женщин с преэклампсией в анамнезе (29%)
по сравнению с контрольной группой (16%, отношение шансов (OR)
).2.0, 95% доверительный интервал (ДИ) 1,2-3,7)
(÷
2
= 9,99, p = 0,0068) при сравнении генотипа Tyr113
Tyr113 с генотипом Tyr113 His113
и His113 His113. Подгруппа
Таблица 1 Характеристики случаев и контроля в период, когда они были госпитализированы по поводу
преэклампсии
Преэклампсия без
HELLP (n = 87)
Преэклампсия с
HELLP (n = 96)
Небеременная
контрольная (n = 151)
Возраст (лет) 28 (19–40) 28 (19–39) 30 (18–42)
Гестационный возраст (нед
дня
)) 31
+5
(26
+3
–38
+2
) 31
+1
(23
+6
–37
+6
+6
+) —
Нерожавшие 67 (79%) 83 (86%) —
Диастолическое АД (мм рт. Ст.) 110 (90–160) 110 (90–145) ND
Значения даны в виде медианы (диапазон) или чисел (в процентах) ) в зависимости от ситуации.
АД, артериальное давление; НД, не определено.
Таблица 2 Распределение полиморфных вариантов в экзонах 3 и 4 гена EPHX
Вариант гена
Преэклампсия
без HELLP
(n = 87)
Преэклампсия
с HELLP
(n == 96)
Преэклампсия, все
(n = 183)
Контрольные
(n = 151)
Экзон 3 Tyr113 Tyr113 27 (31) 24 (25) 51 (28) * 24 (16) *
Tyr113 His113 22 (25) 26 (27) 48 (26) 60 (40)
His113 His113 38 (44) 46 (48) 84 (46) 67 (44)
Экзон 4 His139 His139 54 (62 ) 49 (51) 103 (56) 96 (64)
His139 Arg139 27 (31) 40 (42) 67 (37) 50 (33)
Arg139 Arg139 6 (7) 7 (7) 13 (7) 5 (3)
Оценка генетических полиморфизмов описана в Методах.Проценты указаны в паспорте
тезисов. Статистическая значимость различий между пациентом и контрольной группой оценивалась с помощью критерия хи-квадрат
.